Особенности развития гаметофитов двух видов папоротников российского Дальнего Востока
Главная » Что мы делаем » Полнотекстовые статьи и лекции » Храпко О.В. » Особенности развития гамет…
Храпко О.В., Баранова О.Ю.
Опубликовано в: Интродукционные центры Дальнего Востока России: Итоги исследований (Матер. Первой отчетной сессии регионального Совета ботанических садов Дальнего Востока 10-11 октября 2001 г., Владивосток). Владивосток: Дальнаука, 2002. С. 199-202
Папоротники — довольно древняя и своеобразная группа растений, характеризующаяся сложным циклом развития, состоящим из двух фаз — фазы гаметофита и фазы спорофита. Как гаметофит, так и спорофит представляют собой отдельные индивиды, отличающиеся морфологическими, биологическими особенностями, различными требованиями к экологическим условиям.
Число литературных источников, в которых имеются материалы о гаметофитах папоротников, невелико, в качестве объектов исследований в ряде из них выступают виды, встречающиеся и на российском Дальнем Востоке (Карнишина, 1975; Шорина, 1987а; и др. ). Хотелось бы выделить несколько обобщающих работ данного плана. Так, В.К. Nayar и S. Kaur (1969, 1971) на основе анализа многочисленного материала выделили типы развития нити гаметофита и типы развития пластинки гаметофита гомоспоровых папоротников. Н.И. Шориной (1987б, 1991, 2001) были выделены следующие периоды и возрастные состояния онтогенеза гаметофитов: латентный (покоящиеся споры), виргинильный (проростки), ювенильный (имматурные и молодые гаметофиты), дефинитивный (зрелые) и сенильный (старые гаметофиты). Общие вопросы онтогенеза ряда видов папоротников Южной Сибири были рассмотрены И.И. Гуреевой (2001), предложившей свою периодизацию возрастно-половых состояний гаметофита.
). Хотелось бы выделить несколько обобщающих работ данного плана. Так, В.К. Nayar и S. Kaur (1969, 1971) на основе анализа многочисленного материала выделили типы развития нити гаметофита и типы развития пластинки гаметофита гомоспоровых папоротников. Н.И. Шориной (1987б, 1991, 2001) были выделены следующие периоды и возрастные состояния онтогенеза гаметофитов: латентный (покоящиеся споры), виргинильный (проростки), ювенильный (имматурные и молодые гаметофиты), дефинитивный (зрелые) и сенильный (старые гаметофиты). Общие вопросы онтогенеза ряда видов папоротников Южной Сибири были рассмотрены И.И. Гуреевой (2001), предложившей свою периодизацию возрастно-половых состояний гаметофита.
Целью наших исследований явилось изучение особенностей развития гаметофитов двух видов птеридофлоры российского Дальнего Востока (сем. Athyriaceae Ching: Athyrium sinense Rupr.; сем. Aspidiaceae Mett. et Frank: Dryopteris crassirhizoma Nakai). В статье использованы обобщенные данные наблюдений за развитием гаметофитов папоротников, проведенных в 1994 и 2000 г. Споры собирались в августе названных годов, посевы производились на водный раствор Кнопа в чашки Петри. Подсчитывались проросшие и не проросшие споры в поле зрения микроскопа в 5 повторностях. В последующие периоды наблюдения проводились за стадиями развития гаметофитов, наступление фазы отмечалось по двум датам — появлению первых гаметофитов, вступивших в эту фазу, и по дате, когда насчитывалось максимальное число гаметофитов в данной фазе. Фазы развития зарисовывались. Поскольку наступление фаз в значительной мере зависит от внешних условий (температурный режим) данные о длительности той или иной фазы мы не приводим.
Споры собирались в августе названных годов, посевы производились на водный раствор Кнопа в чашки Петри. Подсчитывались проросшие и не проросшие споры в поле зрения микроскопа в 5 повторностях. В последующие периоды наблюдения проводились за стадиями развития гаметофитов, наступление фазы отмечалось по двум датам — появлению первых гаметофитов, вступивших в эту фазу, и по дате, когда насчитывалось максимальное число гаметофитов в данной фазе. Фазы развития зарисовывались. Поскольку наступление фаз в значительной мере зависит от внешних условий (температурный режим) данные о длительности той или иной фазы мы не приводим.
Споры изучаемых нами видов, как и большинства представителей гомоспоровых папоротников, билатеральные, в боковом положении их очертания бобовидные, в проксимальном — овальные, в экваториальном — округлые (см. рис.). Периспорий спор Athyrium sinense нерельефный, прилегающий к зерну споры, в виде тонкого зернистого слоя. Длина спор в боковом положении 41,2±0,3 мк, ширина -26,3±0,4 мк.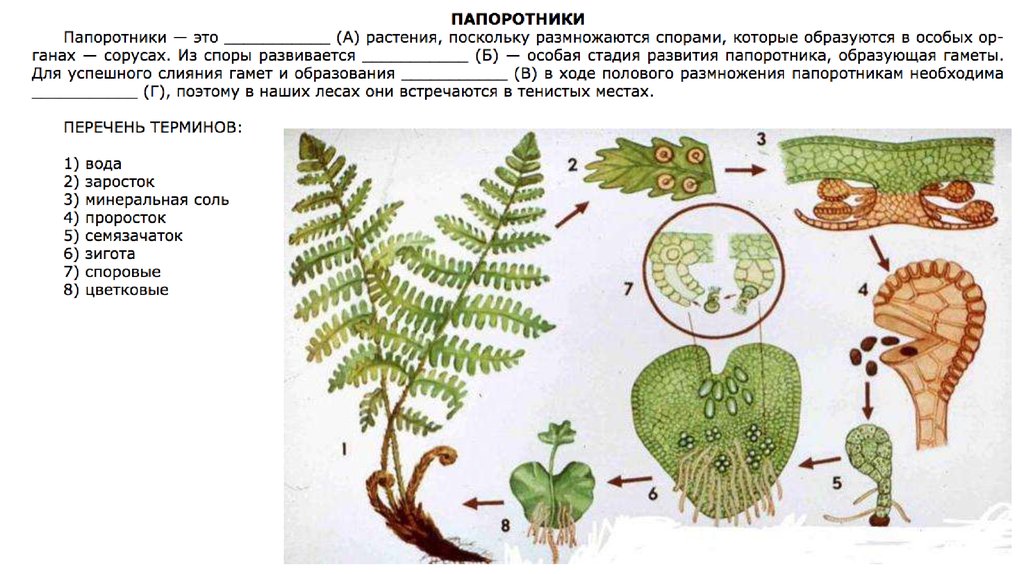 По форме, характеру периспория, размерам они очень сходны со спорами близких видов — Athyrium filix-femina (L.) Roth и A. monomachii (Kom.) Kom. (Кутафьева, 1972; Храпко, 1996). По классификации, приведенной В.П. Гричук и М.Х. Моносзон (1971) споры Athyrium sinense можно отнести к типу VII.
По форме, характеру периспория, размерам они очень сходны со спорами близких видов — Athyrium filix-femina (L.) Roth и A. monomachii (Kom.) Kom. (Кутафьева, 1972; Храпко, 1996). По классификации, приведенной В.П. Гричук и М.Х. Моносзон (1971) споры Athyrium sinense можно отнести к типу VII.
Споры Dryopteris crassirhizoma четко бобовидные, с тонким прозрачным, лишенным скульптуры, складчатым периспорием (см. рис.). По контуру периспорий крупнобугорчатый, образует широкую волнистую кайму различной ширины. Складки недлинные, беспорядочно разбросаны, имеют вид мало изогнутых извилин. Длина спор в боковом положении 46,9±0,9 мк, ширина — 33,1 ±0,4 мк. С учетом морфологических особенностей и характера периспория споры данного вида отнесены к типу 1.2 (Гричук, Моносзон, 1971 как Dryopteris buschiana Fomin).
Для прорастания спор папоротников необходимо сочетание целого ряда благоприятных факторов. Поскольку в естественных местах произрастания условия не всегда оказываются оптимальными, значительная часть высыпавшихся из спорангиев спор не прорастает.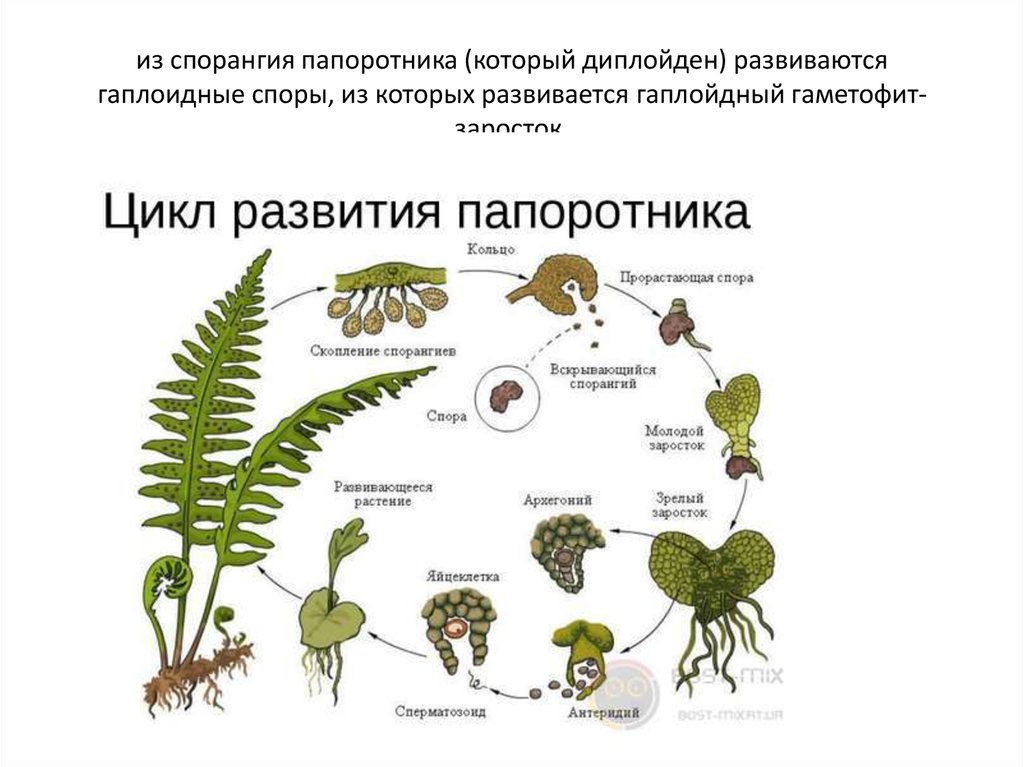 Это компенсируется большим количеством спор, продуцируемых одной особью папоротников. Так, по литературным данным (Schneller, 1975), на одном взрослом растении Dryopteris filix-mas (L.) Schott созревает около 100 млн. спор. Основная масса спор высыпается на протяжении 2-3 недель после созревания, но некоторое их количество остается на вайях в сорусах на протяжении всего зимнего периода. Всхожесть таких спор у изучаемых нами видов составляла от 90 до 80%. При посеве спор после 1,5 года хранения в лабораторных условиях всхожесть спор обоих видов не превышала 10%, при посеве через 4-6 мес. она колебалась от 63 до 98%.
Это компенсируется большим количеством спор, продуцируемых одной особью папоротников. Так, по литературным данным (Schneller, 1975), на одном взрослом растении Dryopteris filix-mas (L.) Schott созревает около 100 млн. спор. Основная масса спор высыпается на протяжении 2-3 недель после созревания, но некоторое их количество остается на вайях в сорусах на протяжении всего зимнего периода. Всхожесть таких спор у изучаемых нами видов составляла от 90 до 80%. При посеве спор после 1,5 года хранения в лабораторных условиях всхожесть спор обоих видов не превышала 10%, при посеве через 4-6 мес. она колебалась от 63 до 98%.
Литературные материалы (Weinberg, Voeller, 1969; Kurichan, Kalarani, 1975; и др.) и наши наблюдения показывают, что необходимым условием для прорастания спор большинства папоротников является свет. Наиболее оптимальны для прорастания Athyrium sinense, Dryopteris crassirhizoma температуры в пределах 20-22°С, кислотность среды должна составлять рН=5,2-6,8 (Otto et al.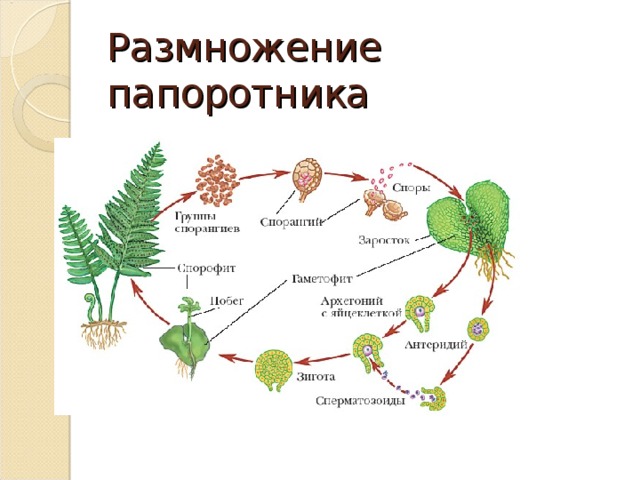 , 1984). При благоприятных условиях прорастание спор изучаемых видов отмечалось через 5-7 дн. после посева.
, 1984). При благоприятных условиях прорастание спор изучаемых видов отмечалось через 5-7 дн. после посева.
Прорастание спор Athyrium sinense и Dryopteris crassirhizoma начинается с появления первой клетки, внутри которой хорошо просматриваются хлоропласты. Эта клетка делится перегородкой, перпендикулярной полярной оси споры, в результате чего образуется первый ризоид и клетка нити гаметофита. Первый ризоид вытягивается параллельно полярной оси. Серией делений терминальной клетки перегородками, параллельными первой, формируется нить гаметофита. Сформировавшаяся нить гаметофита состоит из 5-8 хлорофиллосодержащих клеток и несет 1-2 бесцветных ризоида. На этой стадии у гаметофитов Dryopteris crassirhizoma она заканчивается терминальным волоском. Подобный тип прорастания спор и формирования нити гаметофита названы в литературе (Nayar, Kaur, 1969, 1971) Vittaria-тшюи. Этот тип характерен как для представителей сем. Athyriaceae и Aspidiaceae, так и в целом для класса Filicopsida.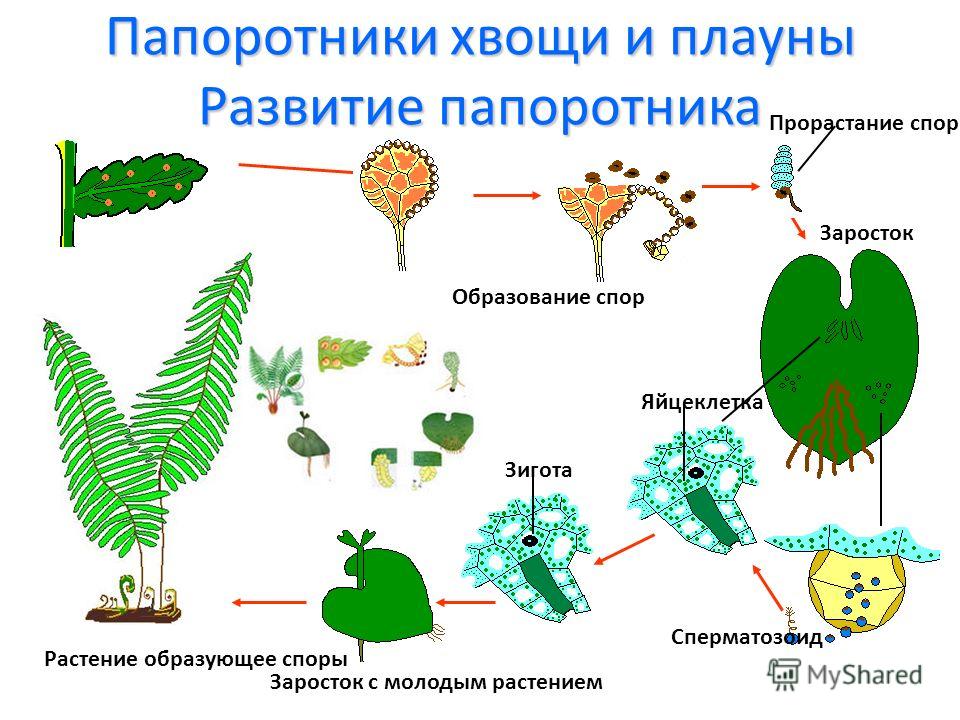
Формирование пластинки гаметофита Athyrium sinense продолжается далее делением терминальной клетки двумя параллельными тангентальными перегородками. Средняя из получившихся в результате такого деления трех клеток дает начало меристеме. В дальнейшем формирование сердцевидного гаметофита идет путем многократного деления дочерних клеток перегородками параллельными и перпендикулярными первой. Меристема располагается в выемке сердцевидной пластинки гаметофита. По характеру формирования пластинки гаметофита данный вид отнесен к Adiantum-типу.
Формирование пластинки гаметофита Dryopteris crassirhizoma происходит по другой схеме. У нити этого вида делится вторая или третья клетки на терминальном конце нити. Одна из образовавшихся при делении клеток дает начало меристеме, в результате последующих многократных делений образуется пластинка гаметофита, которая на ранних этапах развития асимметрична. Уже на этих этапах по краю пластинки заметны секреторные волоски. В дальнейшем гаметофит Dryopteris crassirhizoma приобретал характерную для большинства папоротников форму сердцевидной пластинки с симметричными крыльями. Такой тип развития пластинки гаметофита получил название Aspidium-типа (Nayar, Kaur, 1969, 1971). Зрелый гаметофит данного вида округло-сердцевидный, обильно покрыт секреторными волосками.
Такой тип развития пластинки гаметофита получил название Aspidium-типа (Nayar, Kaur, 1969, 1971). Зрелый гаметофит данного вида округло-сердцевидный, обильно покрыт секреторными волосками.
Рис. Споры и развитие гаметофита Dryopteris crassirhizoma : a — боковое положение споры; б — проксимальное; в — экваториальное положение споры; 1 -прорастание споры; 2 — образование нити гаметофита; 3 — формирование пластинки гаметофита; с — спора; хл — хлоропласты; р — ризоид; вл — секреторный волосок.
Таким образом, изучение онтогенеза гаметофитов Athyrium sinense и Dryopteris crassirhizoma показало, что в развитии гаметофитов данных видов имеются как общие черты, так и различия. Отличия отмечены в строении периспория, но форма и очертания спор обоих видов сходны. Сходны и отнесены к одному (Vittaria-тяп) типу прорастание спор и последовательность формирования нити гаметофитов. Значительные различия имеются в ходе формирования пластинок гаметофитов, у Athyrium sinense оно идет по Adiantum-типу, у Dryopteris crassirhizoma — по Aspidium-типу. Зрелые гаметофиты обоих изучаемых видов сходны по форме и строению. Отличие состоит в том, что пластинка зрелого гаметофита Dryopteris crassirhizoma покрыта секреторными волосками.
Зрелые гаметофиты обоих изучаемых видов сходны по форме и строению. Отличие состоит в том, что пластинка зрелого гаметофита Dryopteris crassirhizoma покрыта секреторными волосками.
Гричук В.П., Моносзон М.Х. Определитель однолучевых спор папоротников из семейства Polypodiaceae R. Br., произрастающих на территории СССР. Новосибирск: Наука, 1971. 125 с.
Гуреева И.И. Равноспоровые папоротники Южной Сибири. Систематика, происхождение, биоморфология, популяционная биология. Томск: Изд-во Томского ун-та, 2001. 158 с.
Карнишина Л.М. Особенности развития и морфологии гаметофитов и юных спорофитов представителей p. Dryopteris Adans. s.l. (на примере видов Московской области) // Природные процессы и ресурсы Подмосковья. М., 1975. С. 132-141.
Кутафьева Т.К. Морфология спор плаунов, хвощей и папоротников Средн. Сибири // Ботан. журн. 1972. Т. 57, № 10. С. 1309-1316.
Храпко О.В. Папоротники хвойно-широколиственных лесов Приморского края (биология, экология, перспективы использования и задачи охраны генофонда). Владивосток: ДВО РАН СССР, 1989. 124 с.
Владивосток: ДВО РАН СССР, 1989. 124 с.
Храпко О.В. Папоротники юга Дальнего Востока России (биология, экология, вопросы охраны генофонда). Владивосток: Дальнаука, 1996. 200 с.
Шорина Н.И. Сравнительная морфология и морфогенез гаметофитов у равноспоровых папоротников // 5 всесоюз. школа по теорет. морфологии растений. 15-18 сент. 1987. Львов, 1987 а. С. 87-92.
Шорина Н.И. Возрастные состояния гаметофитов папоротников и возрастно-половая структура их популяций // Популяционная экология растений: Матер, конф. к 85-летию со дня рожд. А.А. Уранова. 27 янв.-1 февр. 1986 г. М., 1987 б. С. 43-48.
Шорина Н.И. Двойственность популяционной экологии равноспоровых папоротников // Экология популяций. М.: Наука, 1991. С. 180-198.
Шорина Н.И. Популяционная биология гаметофитов равноспоровых Polypodiophyta II Экология. 2001. № 3. С. 182-187.
Kurichan P.I., Kalarani Т.К. Light dependence of spore germination in a few leptosporangiate ferns // New Bot. 1975. Vol. 2, N 2-3. P. 156-157.
P. 156-157.
Nayar B.K., Kaur S. Gametophytes of homosporous ferns // Bot. Rev. 1971. Vol. 37. P. 295-396.
Nayar B.K., Kaur S. Types of prothallial development in homosporous ferns // Phytomorphology, 1969. Vol. 19, N2. P. 179-188.
Otto E. A., Crow J.H., Kirby E.D. Effect of acidic growth conditions on spore germination and reproductive development in Dryopteris marginalis (2) // Ann. Bot. 1984. Vol. 53, N 3. P. 439-442.
Schneller J.J. Untersuchungen an einheimischen Farnen, insbesondere der Dryopteris filix-mas-gruppe. 3. Teil. Okologiche Untersuchungen // Ber. Schweiz. bot. Ges. 1975. 85, N2. S. 110-159.
Weinberg E.S., Voeller B.R. External factors inducing of fern spores // Amer. Fern J. 1969. Vol. 59, N4. P. 153-167.
Собраны геномы двух папоротников — PCR News
В настоящее время существует более 10500 видов папоротников — это самый обширный таксон растений после покрытосеменных. При этом 99% видов относятся к равноспоровым, или гомоспоровым (то есть продуцируют споры, из которых вырастают обоеполые, а не раздельнополые гаметофиты;
пример жизненного цикла такого папоротника). Папоротники — ближайшие родственники семенных растений, поэтому их изучение даст более глубокое понимание эволюции всей наземной флоры. Однако их огромные геномы (в среднем 12,3 Гб; максимум 147 Гб) и большое число хромосом (в среднем 40,5; максимум 720) препятствуют получению высококачественных геномных сборок, а следовательно, и геномным исследованиям этой филогенетически важной группы растений.
Папоротники — ближайшие родственники семенных растений, поэтому их изучение даст более глубокое понимание эволюции всей наземной флоры. Однако их огромные геномы (в среднем 12,3 Гб; максимум 147 Гб) и большое число хромосом (в среднем 40,5; максимум 720) препятствуют получению высококачественных геномных сборок, а следовательно, и геномным исследованиям этой филогенетически важной группы растений.
После многих лет работы две независимые международные группы ученых опубликовали в журнале Nature Plants
Геном цератоптериса — это 7,46 Гб ДНК, упакованной в 39 пар хромосом, что более чем вдвое превышает размер человеческого генома, а геном адиантума — 4,83 ГБ ДНК и 30 пар хромосом.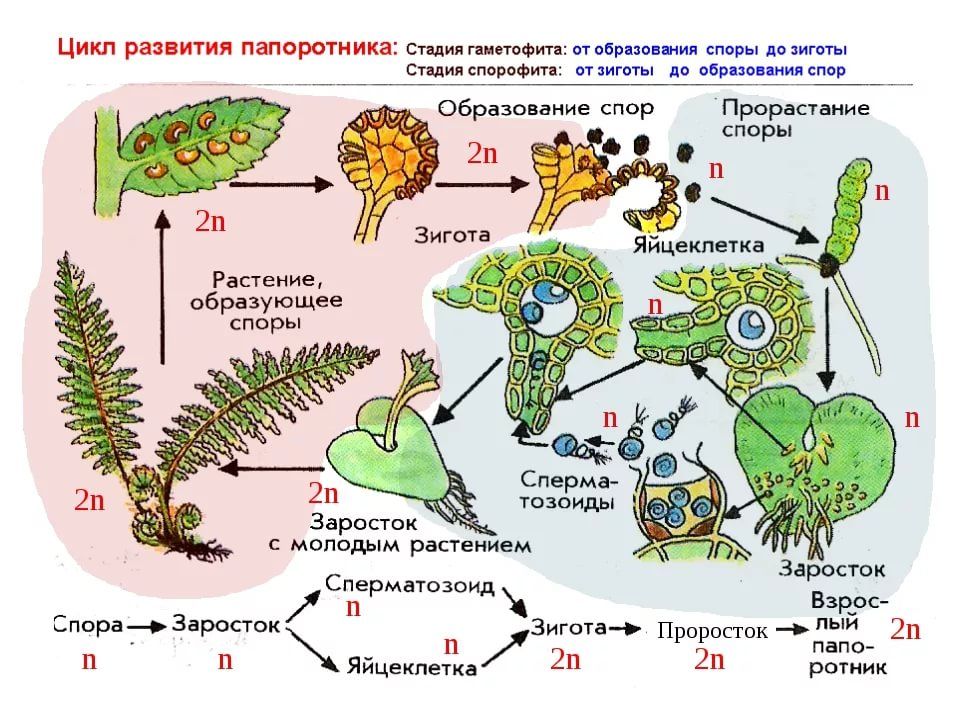 Ранее считалось, что столь большой размер геномов папоротников обусловлен недавней полногеномной дупликацией (whole genome duplication, WGD). Различные копии генов (паралоги) обычно становятся сырьем для формирования новых признаков. Дупликации генома часто встречаются у покрытосеменных растений, в том числе сельскохозяйственных.
Ранее считалось, что столь большой размер геномов папоротников обусловлен недавней полногеномной дупликацией (whole genome duplication, WGD). Различные копии генов (паралоги) обычно становятся сырьем для формирования новых признаков. Дупликации генома часто встречаются у покрытосеменных растений, в том числе сельскохозяйственных.
Исследователи ожидали, что большая часть хромосом цератоптериса и адиантума должна быть идентичной. Чтобы оценить вклад дупликаций, они искали в геномах крупные синтенические блоки генов-паралогов. Однако у цератоптериса было обнаружено только 45 синтенических блоков, включающих 367 генов, у адиантума — 7 блоков и в них 96 генов паралогов, что составляет всего 0,31% от общего числа его генов. Хотя древняя WGD все же имела место у обоих видов, не она внесла вклад в увеличение размеров их геномов; по-видимому, значительная доля полиплоидных участков впоследствии была потеряна.
Также было обнаружено, что 85% генома C. richardii состоит из транспозонов, и такая же пропорция наблюдалась у A. capillus-veneris. Можно сказать, что геномы обоих видов в основном содержат генетический мусор, накопленный за миллионы лет.
capillus-veneris. Можно сказать, что геномы обоих видов в основном содержат генетический мусор, накопленный за миллионы лет.
«Кажется, мы обнаруживаем, что такие растения, как цветковые, геномы которых в среднем намного меньше, чем у папоротников, просто лучше избавляются от ненужной ДНК. Они лучше сбрасывают запасные хромосомы и даже уменьшаются после небольших дупликаций», — сказал Блэйн Маршан (Blaine Marchant), первый автор статьи о цератоптерисе.
Известно, что растительноядные насекомые незначительно повреждают папоротники. Анализ цератоптериса выявил несколько генов — гомологов Tma12, кодирующего инсектицидный белок. Этот ген ранее находили и в геномах других папоротников. Цератоптерис, по-видимому, получил его от бактерий посредством горизонтального переноса. Кроме того, авторы обнаружили множественные копии гена порообразующего токсина, подобного бактериальному аэролизину: он делает отверстие в мембране клетки, что приводит к ее гибели. Учитывая, что несколько копий гена были обнаружены в трех отдельных хромосомах, вполне вероятно, что перенос происходил более одного раза.
Учитывая, что несколько копий гена были обнаружены в трех отдельных хромосомах, вполне вероятно, что перенос происходил более одного раза.
Особый интерес представляет исследование эволюционного перехода от бессемянных растений к семенным. Анализ генов цератоптериса, гомологичных тем, которые имеют решающее значение для индукции цветения и архитектуры семенных растений, показал, что у папоротника некоторые из них, по-видимому, связаны с развитием спор.
Авторы второй статьи, которые исследовали адиантум, проанализировали транскриптомы, полученные из разных тканей спорангиев на разных стадиях развития папоротника. Паттерны экспрессии генов в спорангиях проявили сходство с экзиной цветковых растений (наружная утолщенная оболочка пыльцевого зерна). Сходным оказался и молекулярный механизм LEC1 -опосредованного накопления запасных белков у A.capillus-veneris и Arabidopsis.
Таким образом, гены, управляющие развитием семян, могли произойти от генов, контролирующих развитие спорангиев папоротников.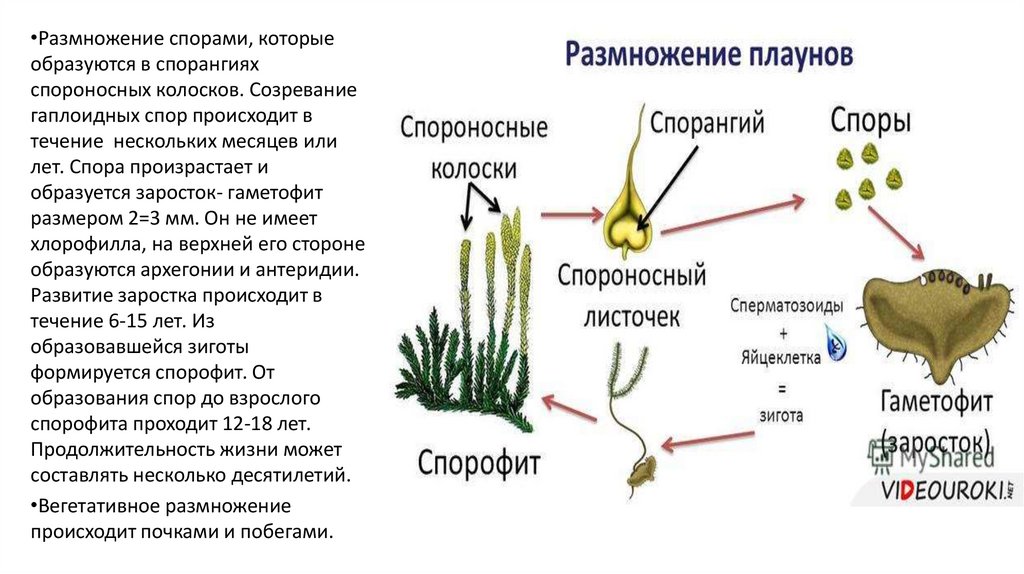
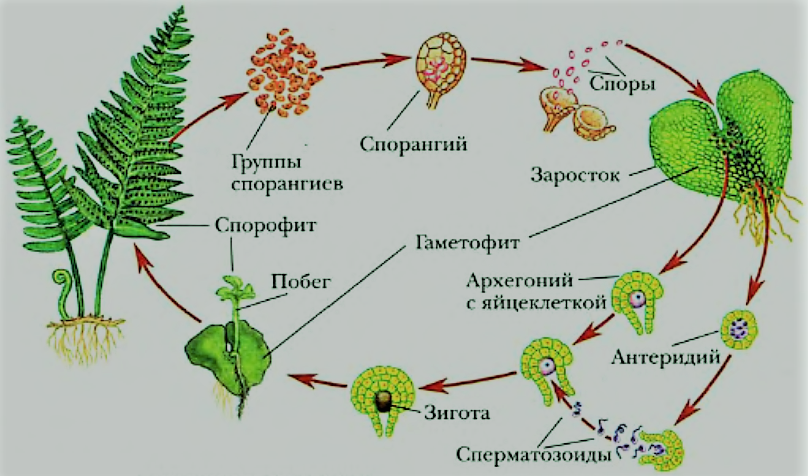
Размножение папоротника
Размножение спорами
Папоротники и хвощи имеют два свободноживущих поколения:
- диплоидное спорофитное поколение (спорообразующее растение) и
- гаплоидный гаметофит поколение (растение, производящее гаметы).
Растения, которые мы видим как папоротники или хвощи, относятся к поколению спорофитов. Летом спорофит обычно выпускает споры. Споры должны приземлиться на подходящую поверхность, например, на влажную защищенную зону, чтобы прорасти и превратиться в гаметофиты.
Зрелый гаметофит многих наших папоротников выглядит как маленькое плоское зеленое сердце размером с ноготь. Мужские и женские репродуктивные структуры развиваются на нижней поверхности одного и того же, а чаще на разных гаметофитных растениях. При половой зрелости мужские структуры выделяют сперму, которая проплывает через пленку воды во влажной среде обитания, чтобы оплодотворить яйцеклетку в женской структуре.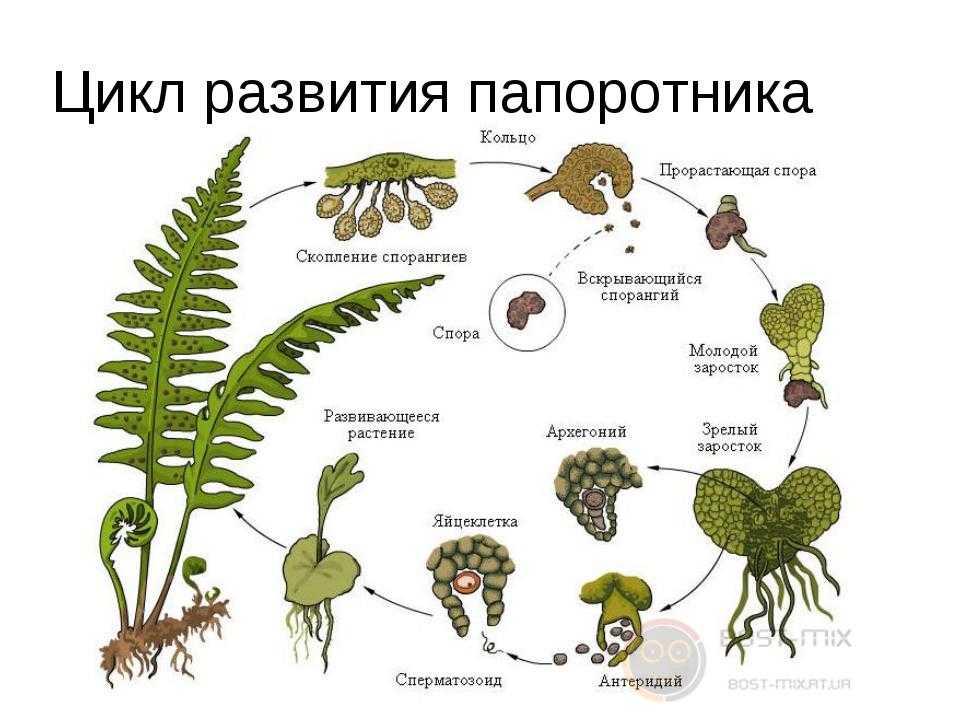
Многие гаметофиты обычно растут в непосредственной близости друг от друга, и у большинства папоротников и хвощей сперма одного гаметофита, скорее всего, оплодотворит яйцеклетку другого гаметофита. Это оплодотворенное яйцо развивается в зародыш, который является началом поколения диплоидного спорофита. Первый крошечный лист спорофита выходит из нижней части гаметофита (который распадается), растет вверх и вскоре становится растением, которое мы видим в виде папоротника или хвоща.
Этот зрелый хрупкий гаметофит папоротника имеет диаметр около ½ дюйма и растет на почве, которая почти постоянно влажная.
Части гаметофита.
Вы можете выращивать папоротники из спор. Американское общество папоротников описывает, как собирать споры и выращивать папоротники из спор. Вы также можете получить споры самых разных папоротников от Американского общества папоротников.
Вегетативное размножение
Луковицы и папоротники
Луковицы представляют собой небольшие вегетативные почки, которые растут на ветвях нескольких видов папоротников.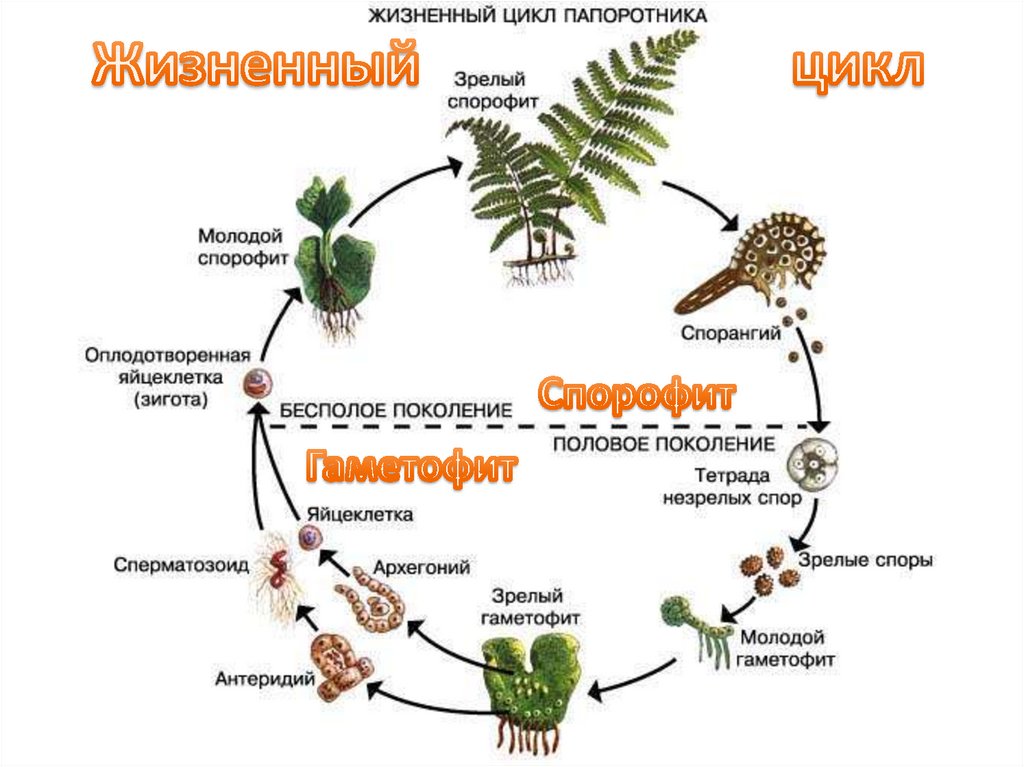 Когда луковица прижимается к земле или падает с папоротника, она может укорениться и превратиться в зрелый папоротник. Примеры папоротников, которые могут размножаться луковицами, — 9.0005
Когда луковица прижимается к земле или падает с папоротника, она может укорениться и превратиться в зрелый папоротник. Примеры папоротников, которые могут размножаться луковицами, — 9.0005
Остролистный папоротник Андерсона на северо-западном побережье производит луковицу к кончику ветви. При прижатии к земле луковицы вырастают в новые растения; поэтому этот папоротник часто растет колониями растений разного возраста, как показано на этой фотографии колонии остролистного папоротника Андерсона.
Ходячий папоротник получил свое название из-за интересного способа создания новых растений. Его простые листья имеют длинные заостренные кончики. Когда эти кончики касаются земли, развивается новый папоротник; таким образом, папоротник «гуляет» по ландшафту.
Колония остролистника Андерсона ( Polystichum andersonii ).
Остролистник Андерсона ( Polystichum andersonii ). Красная стрелка указывает на луковицу, расположенную ближе к кончику вайи.
Остролистник Андерсона ( Polystichum andersonii ). Луковица превращается в новый папоротник. Красная стрелка указывает на крошечную ветвь папоротника, выходящую из луковицы.
Луковица превращается в новый папоротник. Красная стрелка указывает на крошечную ветвь папоротника, выходящую из луковицы.
Рост корневища (стебля)
Папоротники с ползучим корневищем распространяются по мере роста корневища выше или ниже субстрата. Корни и листья образуются возле кончика удлиняющегося и ветвящегося корневища. По мере роста растения корневище может разрываться, отделяя сегменты папоротника. Отделившийся сегмент живет, потому что корневище укоренено по всей длине и несет листья. Один папоротник может распространиться на большую колонию папоротников. Таким образом распространяются многие папоротники, в том числе дубовый папоротник, буковый папоротник, страусиный папоротник, папоротник-орляк и хвощ обыкновенный.
Выращивание мечевидных папоротников из спор в темные дни зимы
Споры остролистного папоротника (Polystichum) прорастают в специальных контейнерах при контролируемых условиях освещения, температуры и влажности.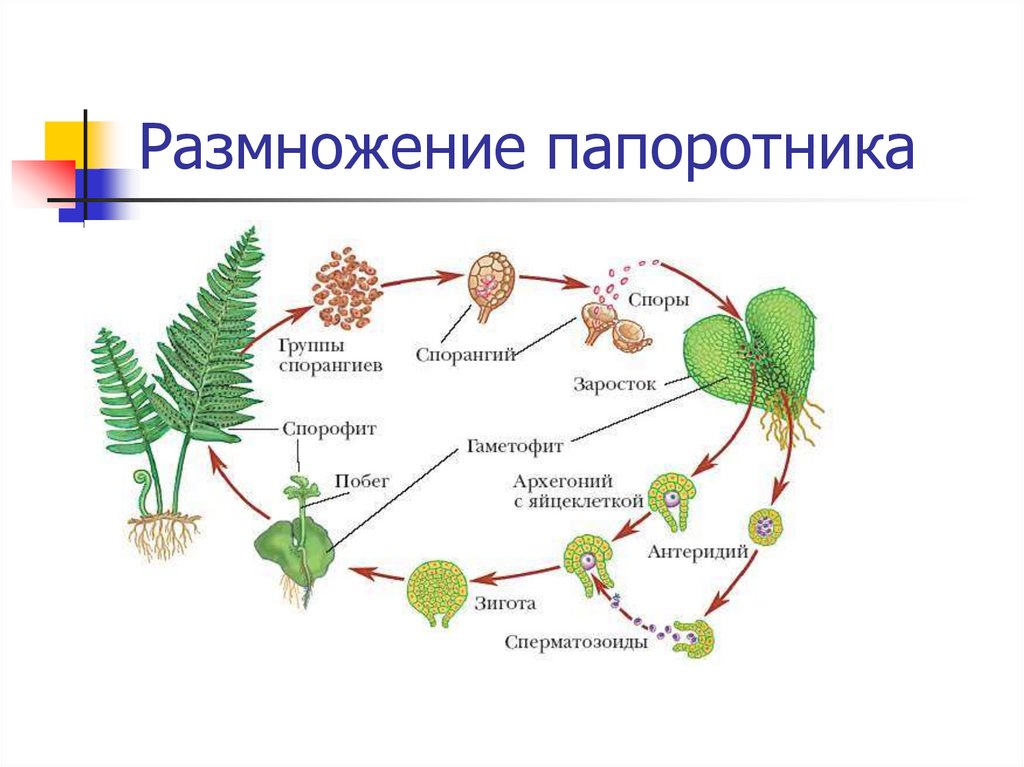
Район рейнджеров Ситка (Национальный лес Тонгасс), Служба национальных парков, Кооперативная служба распространения знаний Университета Аляски. и Геологическая служба США сотрудничает в выращивании мечевидных папоротников из спор в рамках проекта по размножению местных растений. Папоротники-мечи — эффектные папоротники, ценные для озеленения или восстановления растительности.
Четыре вида мечевидных папоротников произрастают на острове Бараноф, где расположена Ситка, и хорошо адаптированы к различным средам обитания и хорошо пересаживаются. Ни один из этих больших вечнозеленых папоротников не является достаточно обильным, чтобы поддерживать коммерческий урожай. Размножение спорами может быть способом получения достаточного количества растений для реставрационных проектов или использования местными землевладельцами. Простые протоколы тестируются для выращивания папоротников. Споры собирали в дикой природе, высевали на стерильную почву в специальные крытые контейнеры. Их обеспечивают светом 12 часов в сутки и тщательно опрыскивают для увлажнения.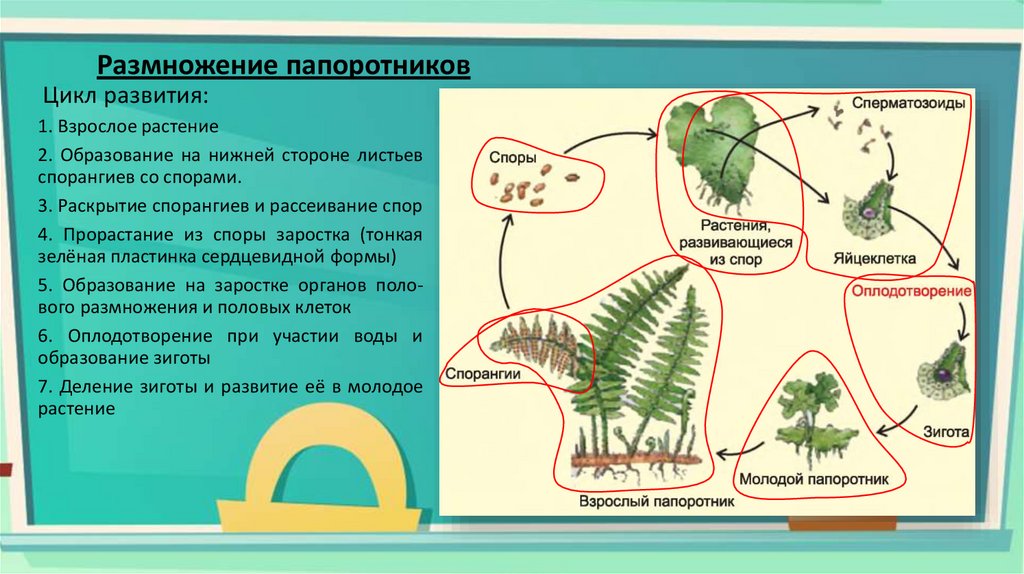
Подробнее (PDF, 152 КБ)…
Жизненный цикл папоротников — Центр научного обучения
Жизненный цикл папоротников отличается от жизненного цикла других наземных растений, поскольку фазы гаметофита и спорофита являются свободными. Этот интерактив иллюстрирует смену поколений папоротников.
Жизненный цикл папоротников отличается от других наземных растений, поскольку и гаметофит, и спорофит живут свободно. Этот интерактив иллюстрирует смену поколений папоротников.
Стенограмма
Мейоз
Мейоз — это тип клеточного деления, при котором образуются гаметы — клетки, содержащие вдвое меньше хромосом, чем родительская клетка. У папоротников такими клетками являются споры.
Стенограмма
Д-р Леон Перри
Типичное большое растение папоротника в результате мейоза производит споры, а число хромосом вдвое меньше, чем у большого родительского растения.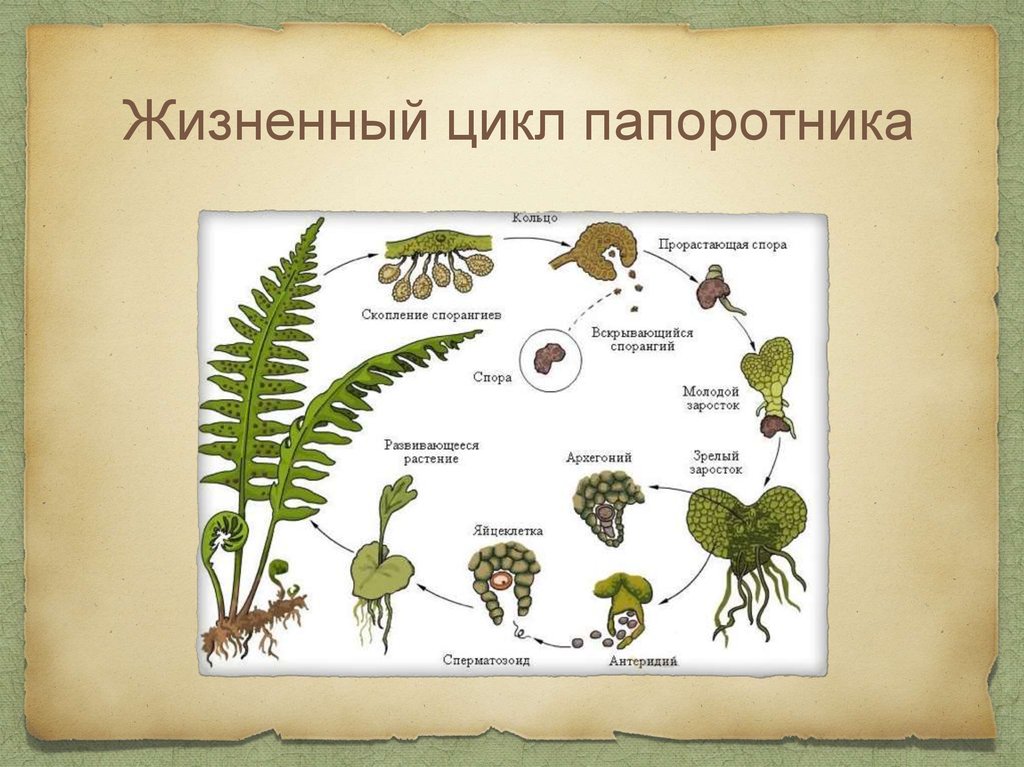
Видео: © University of Waikato
Spores
Споры представляют собой небольшие репродуктивные структуры, выделяющиеся из спорангиев.
Стенограмма
Доктор Леон Перри
Итак, спора является продуктом мейоза. У папоротников он крошечный. Если вы возьмете линейку, подумаете, насколько велик сантиметр, и разделите его на 10, тогда у вас получится миллиметр, а затем снова разделите его на 10, а затем разделите его пополам. Это примерно размер споры.
Видео: © University of Waikato
Микрофотография споры, Австралазийский атлас пыльцы и спор, Creative Commons 3.0
Прорастание спор
Высвобожденные споры вырастают в гаметофиты – очень маленькие структуры сердцевидной формы.
Стенограмма
Доктор Леон Перри
Споры разлетаются по ветру. Если эти споры попадут в подходящее место, они вырастут в то, что называется гаметофитом, то есть в отдельное отдельное растение. Он очень крошечный, может быть, размером с ноготь, и он похож на маленькую, тонкую, маленькую зеленую тарелку.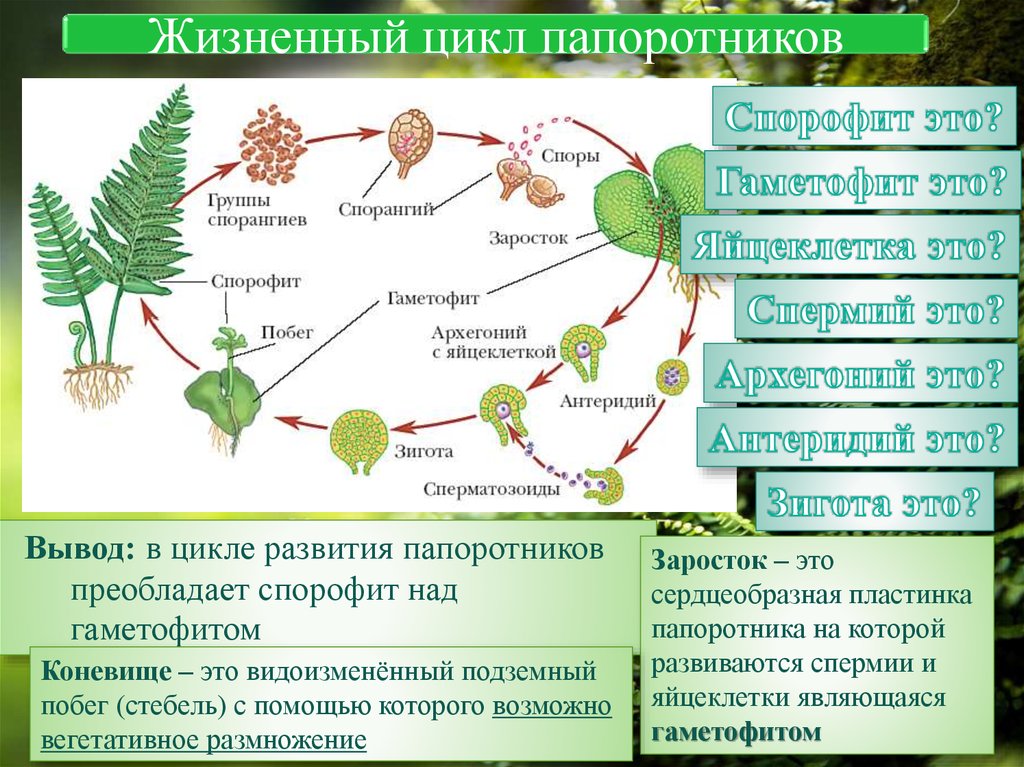 Что это делает, так это производит половые клетки – яйцеклетки и сперму.
Что это делает, так это производит половые клетки – яйцеклетки и сперму.
Видео: © University of Waikato
Микрофотография споры, Australasian Pollen & Spore Atlas, Creative Commons 3.0
Гаметофит (проталлус)
Гаметофиты содержат как женские (архегонии), так и мужские (антеридии) половые органы. Они созревают в разное время, чтобы увеличить шансы перекрестного оплодотворения и генетической изменчивости.
Заросток – гаметофит папоротника. Это зеленая фотосинтетическая структура толщиной в одну клетку, обычно сердцевидной или почковидной формы, длиной 3–10 мм и шириной 2–8 мм. Его очень трудно найти в кустах, так как он такой крошечный. У него нет корней, стеблей или листьев, но есть ризоиды, которые прикрепляют его к почве и помогают всасыванию. На нижней стороне заростка образуются гаметы мужских и женских половых органов. Проталлус имеет как мужские, так и женские половые органы.
Признание: общественное достояние
Архегоний
Архегоний — женский половой орган.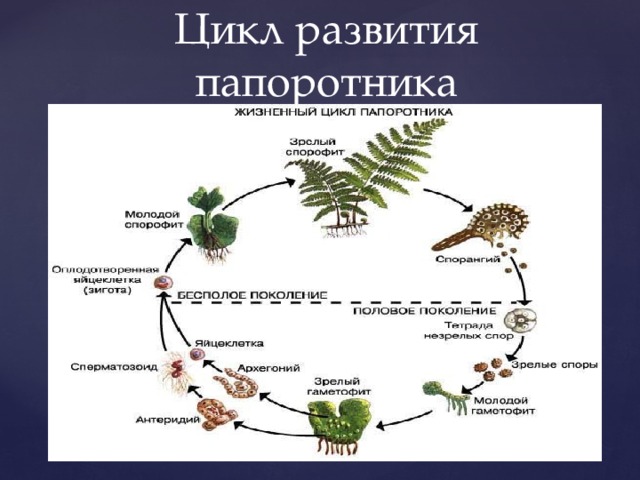 Это колбообразные структуры, которые производят яйцеклетку, до которой добирается сперма, плывущая по шее.
Это колбообразные структуры, которые производят яйцеклетку, до которой добирается сперма, плывущая по шее.
Подтверждение: Университет штата Огайо
Антеридиум
Антеридиум — мужской половой орган. Это небольшие сферические структуры, производящие жгутиковые сперматозоиды.
Благодарность: Университет штата Огайо
Оплодотворение
Оплодотворение происходит, когда яйцеклетка и сперма папоротника объединяются, образуя зиготу. Папоротникам требуется вода, чтобы сперматозоиды могли добраться до яйцеклетки. Зигота представляет собой комбинацию генетического материала яйцеклетки и спермы и содержит полный набор ДНК для формирования нового растения папоротника.
СтенограммаДоктор Леон Перри
Сперматозоидам нужно плыть по воде, чтобы добраться до яйцеклеток. Яйца размещаются или сохраняются в гаметофите, и эта зависимость от воды является причиной того, что папоротники так часто связаны с влажной средой обитания.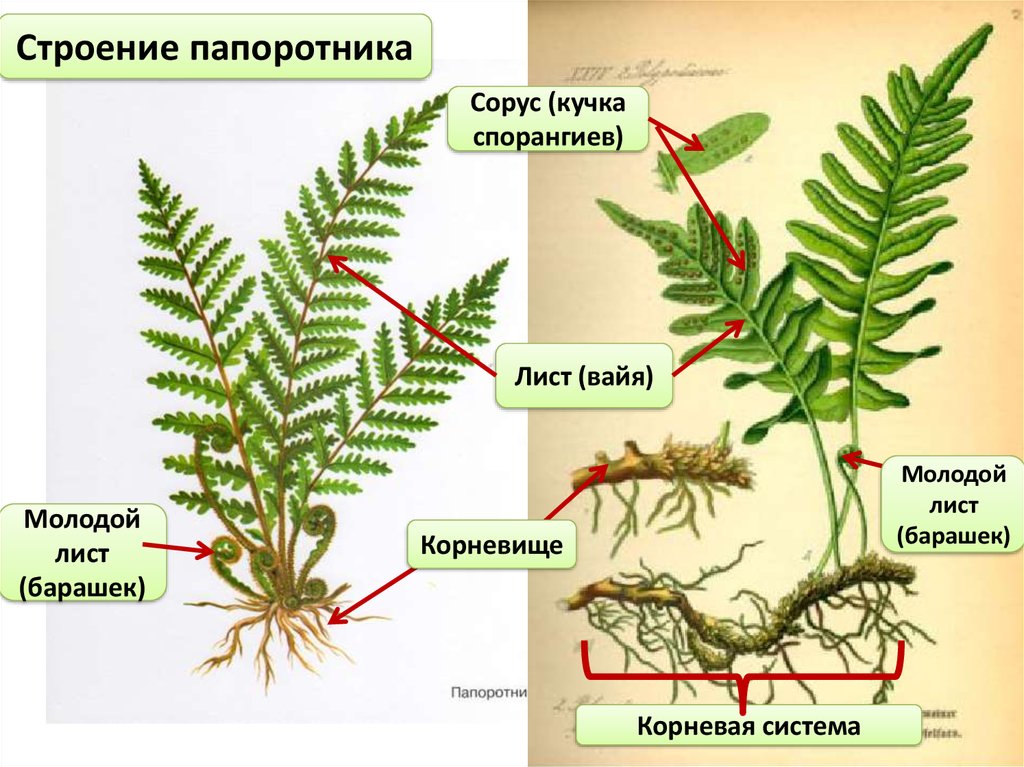 Если сперматозоиду все-таки удается добраться до яйцеклетки, происходит оплодотворение, и именно здесь двое — сперматозоид и яйцеклетка — объединяются. Это удваивает число хромосом и снова дает начало совершенно новому типичному папоротнику, и цикл повторяется.
Если сперматозоиду все-таки удается добраться до яйцеклетки, происходит оплодотворение, и именно здесь двое — сперматозоид и яйцеклетка — объединяются. Это удваивает число хромосом и снова дает начало совершенно новому типичному папоротнику, и цикл повторяется.
Видео: © University of Waikato
Изображение архегония предоставлено Университетом штата Огайо
Развивающийся спорофит
Зигота развивается из заростка (гаметофит папоротника). Он растет с помощью митоза и развивается в молодое растение папоротника.
Благодарность: Джордж Шепард
Зрелый спорофит
Зрелое растение папоротника состоит из трех основных частей – корневища, листьев и спорангиев. Зрелое растение папоротника представляет собой спорофитную структуру, производящую споры, которые высвобождаются из спорангиев.
Благодарность: Стив Эттвуд
Спорангии (сгруппированные в сорусы)
Сорусы представляют собой скопления спорангиев, которые содержат репродуктивные споры.