Практическое занятие «Строение кукушкина льна и сфагнума Строение кукушкина льна и сфагнума», ФГОС
Практическое занятие
По материалам журнала « Биология в школе»
2006-2009год
Строение кукушкина льна и сфагнума
Материал и оборудование: гербарные экземпляры кукушкина льна и сфагнума,
предметное и покровное стекла, пинцет, препаровальная игла, стакан с водой, пипетка, салфетка, микроскоп, лупа.
Ход работы:
Рассмотрите рисунок. Какое строение имеет кукушкин лен? Чем отличаются мужские и женские растения?
Рассмотрите гербарные образцы кукушкина льна. Найдите стебель, листья, ризоиды.
На гербарных экземплярах найдите мужские и женские растения. Чем они отличаются? Обратите внимание на внешнее различие листьев на мужских растениях. Среди обычных узких зеленых листьев имеются розетки более крупных желтоватых листьев. На женских растениях образуется коробочка на длинной ножке.
Среди обычных узких зеленых листьев имеются розетки более крупных желтоватых листьев. На женских растениях образуется коробочка на длинной ножке.
Рассмотрите коробочку с крышечкой на длинной ножке. Какое значение имеет коробочка?
Зарисуйте внешний вид зеленого мха кукушкин лен, обозначьте листья, стебель, ризоиды, ножку, коробочку, колпачок.
Рассмотрите внешнее строение сфагнума. Найдите стебель, различные типы боковых веточек (поникающие и горизонтальные), листья. Есть ли у сфагнума ризоиды?
Приготовьте микропрепарат листа сфагнума. Возьмите предметное стекло, пипеткой капните воду. При помощи пинцета или препаровальной иглы аккуратно оторвите лист, положите его в каплю воды на предметное стекло. Накройте покровным стеклом. Рассмотрите готовый препарат сначала при малом, затем при большом увеличении микроскопа. Найдите узкие длинные хлорофиллоносные клетки и крупные водоносные клетки.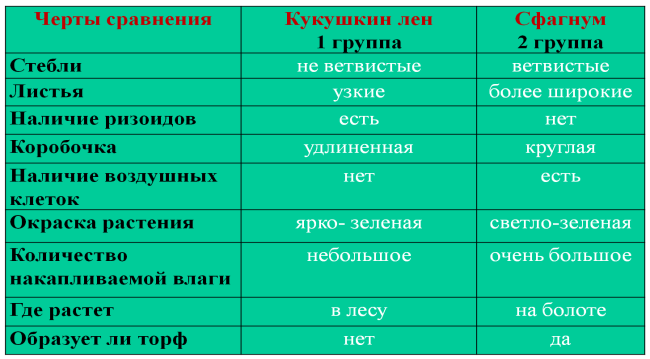 В хлорофиллоносных клетках рассмотрите хлоропласты, а в водоносных клетках — утолщения и поры.
В хлорофиллоносных клетках рассмотрите хлоропласты, а в водоносных клетках — утолщения и поры.
Запишите вывод в тетрадь. Какое строение имеет кукушкин лен и сфагнум? В чем их сходство и отличие? Из каких клеток состоит лист сфагнума? Какую функцию они выполняют?
Строение кукушкина льна и сфагнума
Кукушкин лен — особенности строения, общая характеристика и стадии развития мха
Зеленый мох Кукушкин лён обыкновенный, или Политрихум обыкновенный, вид мхов из рода Кукушкин лен. Широко распространен в нашей стране.
Многолетнее растение, обитающее во влажных местах. Относится к листостебельным мхам. Это один из немногих видов мхов, получивших собственное название в русском языке.
Мох кукушкин лен — происхождение названия
Изящные стебельки кукушкина льна покрыты мелкими темно-зелеными листьями, и выглядит он как лён-долгунец.
На женских растениях появляются коробочки, внешне схожие с кукушкой. Отсюда в русском языке появляется название «кукушкин лён».
Особенности строения
Основные части тела мха это стебель, листья и ризоиды. Коричневатый неветвящийся стебель несет многочисленные линейно-шиловидные листья.
В высоту достигает 30 см. Корни отсутствуют. В нижней части стебля имеются выросты покровной ткани – ризоиды, выполнящиет всасывающую функцию и закрепляющие растение на субстрате. Внешнее покрытие стебля и листьев – эпидерма.
У растения имеется примитивная проводящая система, позволяющая доставлять воду к листьям и транспортировать продукты фотосинтеза по всему организму.
Фотосинтез осуществляется в листьях при помощи хлорофилла в ассимилирующей ткани. Строение листа можно подробно изучить при помощи микроскопа.
Жизненный цикл мха
Последовательность жизненного цикла кукушкина льна схожа с другими представителями класса настоящих мхов. У мхов, в том числе и у кукушкина льна, в жизненном цикле преобладает гаметофит с гаплоидным набором хромосом.
У мхов, в том числе и у кукушкина льна, в жизненном цикле преобладает гаметофит с гаплоидным набором хромосом.
Гаметофит облиственный, в его зеленых листьях происходит фотосинтез, идет транспортировка веществ. На нем развиваются половые клетки — гаметы, после слияния которых образуется организм с диплоидным набором хромосом – спорофит. Рост гаметофита прекращается тогда, когда на нем развивается спорофит.
Спорофит недолговечен и паразитирует на гаметофите. Хлорофилл в его клетках отсутствует, все питательные вещества поступают к нему от гаметофита. Спорофит представляет собой коробочку на ножке.
Коробочка (спорангий) состоит из крышечки и урночки, в которой развиваются споры. Внутри коробочки сеть тонкая пленка – эпифрагма. Она закрывает вход в урночку.
По краю коробочки расположены зубцы – перистом. При изменении влажности зубцы перистома изгибаются внутрь коробочки или наружу, способствуя разбрасыванию спор.
Жизненный цикл развития мха кукушкин лен схематично показан на рисунке.
Размножение мха
Кукушкин лён — двудомное растение, то есть женские и мужские половые клетки развиваются на разных растениях. В женских половых органах (архегониях) появляются яйцеклетки. В антеридиях (мужских половых органах) образуются сперматозоиды.
Мужские растения определяют по темно-коричневым листьям, окружающим антеридий. С каплей воды сперматозоиды попадают к яйцеклетке и оплодотворяют её. Из образовавшейся зиготы прорастает спорофит – тот самый стебелек с коробочкой наверху.
При созревании спор коробочка открывается и рассеивает споры. Споры мелкие, видны только под лупой. Попав в благоприятные условия, спора прорастает в протонему — ветвистую зеленую нить, дающую начало новым растениям – гаметофитам.
Вегетативное размножение мха происходит частями побега.
Распространение и среда обитания
Ареал мха достаточно широк: Северная и Южная Америка, Азия, Европа, Кавказ, Африка, Австралия, Новая Зеландия.
В России произрастает по всей территории от европейской части до Дальнего востока и Сибири, преимущественно в умеренном и арктическом поясах.
Кукушкин лён предпочитает влажную почву и селится в еловых лесах, в низинах, на болотах, сырых лугах. Светолюбивое растение, более высокие соседи сильно угнетают его рост.
Зато, при отсутствии конкуренции за свет, мох агрессивно захватывает большие территории, образуя густые, плотные дернины. Такие дернины не позволяют прорастать семенам других растений и задерживают воду, вызывая заболачивание.
Применение мха кукушкин лен
Используется в следующих сферах человеческой жизни:
-
В медицине.
 Обладает слабительным эффектом. Применяют для остановки кровотечений, как противокашлевое средство, при бронхитах и воспалении легких. Настой травы употребляют при водянке, отеках, запорах. Настой спор обладает противовоспалительным, отхаркивающим, обволакивающим действием.
Обладает слабительным эффектом. Применяют для остановки кровотечений, как противокашлевое средство, при бронхитах и воспалении легких. Настой травы употребляют при водянке, отеках, запорах. Настой спор обладает противовоспалительным, отхаркивающим, обволакивающим действием.
-
В строительстве. Утеплитель для проконопачивания стен. Мох издавна укладывали между бревнами в венцах строящейся избы.
-
В косметологии. Отвар используется в качестве полоскания, укрепляющего волосы и ускоряющего их рост.
Кукушкин лен — особенности строения, общая характеристика и стадии развития мха
Зеленый мох Кукушкин лён обыкновенный, или Политрихум обыкновенный, вид мхов из рода Кукушкин лен. Широко распространен в нашей стране.
Широко распространен в нашей стране.
Многолетнее растение, обитающее во влажных местах. Относится к листостебельным мхам. Это один из немногих видов мхов, получивших собственное название в русском языке.
Мох кукушкин лен — происхождение названия
Изящные стебельки кукушкина льна покрыты мелкими темно-зелеными листьями, и выглядит он как лён-долгунец.
На женских растениях появляются коробочки, внешне схожие с кукушкой. Отсюда в русском языке появляется название «кукушкин лён».
Особенности строения
Основные части тела мха это стебель, листья и ризоиды. Коричневатый неветвящийся стебель несет многочисленные линейно-шиловидные листья.
В высоту достигает 30 см. Корни отсутствуют. В нижней части стебля имеются выросты покровной ткани – ризоиды, выполнящиет всасывающую функцию и закрепляющие растение на субстрате. Внешнее покрытие стебля и листьев – эпидерма.
У растения имеется примитивная проводящая система, позволяющая доставлять воду к листьям и транспортировать продукты фотосинтеза по всему организму.
Фотосинтез осуществляется в листьях при помощи хлорофилла в ассимилирующей ткани. Строение листа можно подробно изучить при помощи микроскопа.
Жизненный цикл мха
Последовательность жизненного цикла кукушкина льна схожа с другими представителями класса настоящих мхов. У мхов, в том числе и у кукушкина льна, в жизненном цикле преобладает гаметофит с гаплоидным набором хромосом.
Гаметофит облиственный, в его зеленых листьях происходит фотосинтез, идет транспортировка веществ. На нем развиваются половые клетки — гаметы, после слияния которых образуется организм с диплоидным набором хромосом – спорофит. Рост гаметофита прекращается тогда, когда на нем развивается спорофит.
Спорофит недолговечен и паразитирует на гаметофите. Хлорофилл в его клетках отсутствует, все питательные вещества поступают к нему от гаметофита. Спорофит представляет собой коробочку на ножке.
Спорофит представляет собой коробочку на ножке.
Коробочка (спорангий) состоит из крышечки и урночки, в которой развиваются споры. Внутри коробочки сеть тонкая пленка – эпифрагма. Она закрывает вход в урночку.
По краю коробочки расположены зубцы – перистом. При изменении влажности зубцы перистома изгибаются внутрь коробочки или наружу, способствуя разбрасыванию спор.
Жизненный цикл развития мха кукушкин лен схематично показан на рисунке.
Размножение мха
Кукушкин лён — двудомное растение, то есть женские и мужские половые клетки развиваются на разных растениях. В женских половых органах (архегониях) появляются яйцеклетки. В антеридиях (мужских половых органах) образуются сперматозоиды.
Мужские растения определяют по темно-коричневым листьям, окружающим антеридий. С каплей воды сперматозоиды попадают к яйцеклетке и оплодотворяют её. Из образовавшейся зиготы прорастает спорофит – тот самый стебелек с коробочкой наверху.
При созревании спор коробочка открывается и рассеивает споры. Споры мелкие, видны только под лупой. Попав в благоприятные условия, спора прорастает в протонему — ветвистую зеленую нить, дающую начало новым растениям – гаметофитам.
Вегетативное размножение мха происходит частями побега. За небольшой промежуток времени в благоприятных условиях растение занимает внушительные площади.
Распространение и среда обитания
Ареал мха достаточно широк: Северная и Южная Америка, Азия, Европа, Кавказ, Африка, Австралия, Новая Зеландия.
В России произрастает по всей территории от европейской части до Дальнего востока и Сибири, преимущественно в умеренном и арктическом поясах.
Кукушкин лён предпочитает влажную почву и селится в еловых лесах, в низинах, на болотах, сырых лугах. Светолюбивое растение, более высокие соседи сильно угнетают его рост.
Зато, при отсутствии конкуренции за свет, мох агрессивно захватывает большие территории, образуя густые, плотные дернины.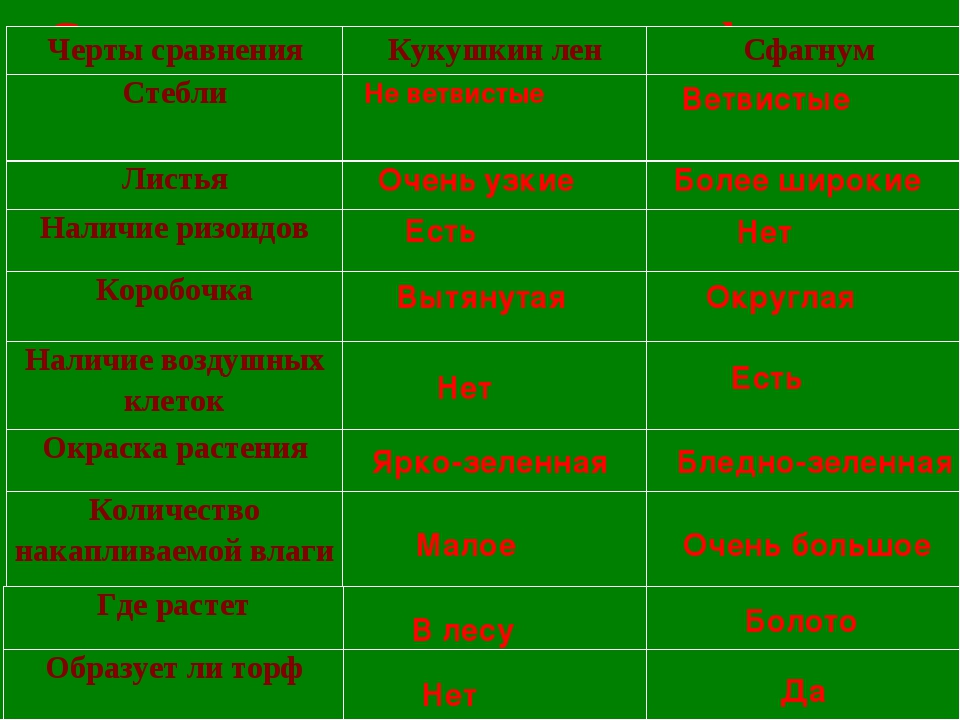
Применение мха кукушкин лен
Используется в следующих сферах человеческой жизни:
В медицине. Обладает слабительным эффектом. Применяют для остановки кровотечений, как противокашлевое средство, при бронхитах и воспалении легких. Настой травы употребляют при водянке, отеках, запорах. Настой спор обладает противовоспалительным, отхаркивающим, обволакивающим действием.
В строительстве. Утеплитель для проконопачивания стен. Мох издавна укладывали между бревнами в венцах строящейся избы.
В косметологии. Отвар используется в качестве полоскания, укрепляющего волосы и ускоряющего их рост.
Предыдущая
БиологияПринцип комплементарности — основа, суть и роль правила в биологии
СледующаяБиологияПродуценты, консументы, редуценты — функции, примеры и роль в экосистеме
Строение кукушкина льна
☰
Кукушкин лен — это род мхов. Обычно при рассмотрении строения кукушкина льна рассматривают один из его видов (кукушкин лен обыкновенный), широко распространенные в России. Это растение растет в местах с повышенной влажностью, нуждается в хорошем освещении. Кукушкин лен образует плотный покров на земле, чем препятствует росту других растений и испарению влаги. В результате может приводить к заболачиванию почв.
Обычно при рассмотрении строения кукушкина льна рассматривают один из его видов (кукушкин лен обыкновенный), широко распространенные в России. Это растение растет в местах с повышенной влажностью, нуждается в хорошем освещении. Кукушкин лен образует плотный покров на земле, чем препятствует росту других растений и испарению влаги. В результате может приводить к заболачиванию почв.
По внешнему виду кукушкин лен представляет собой травянистое растение высотой около 20 см. Стебель обычно не ветвится, имеет зеленовато-бурый оттенок. На стебле располагаются множество зауженных тонких листьев. На них можно увидеть жилку. К почве кукушкин лен прикрепляется с помощью ризоидов (подобия корней, так как у них нет характерных для корней тканей).
Кукушкин лен обыкновенныйСтебель служит для опоры и транспорта веществ. У кукушкина льна есть примитивные проводящие ткани. Так по одним клеткам идет вода с минеральными веществами, по другим — органические вещества.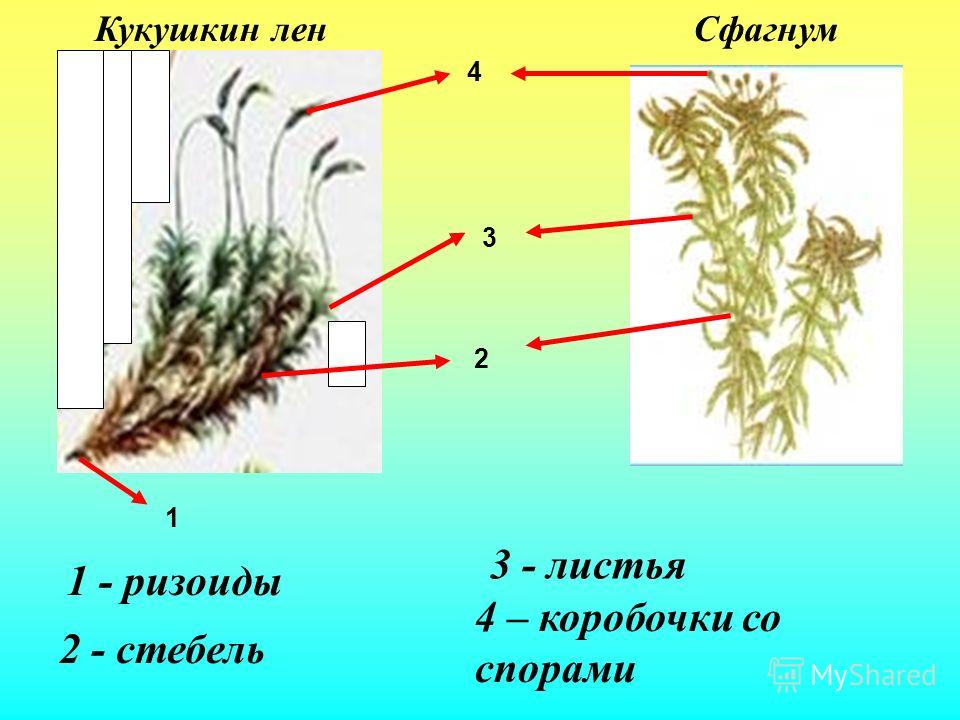
В листьях находятся ряды клеток, основная функция которых — это фотосинтез, т. е. синтез органических веществ. Однако видимо листья также могут поглощать воду.
Ризоиды не только прикрепляют растение к почве, но и всасывают воду с растворенными в ней минеральными веществами.
Кукушкин лен размножается спорами. Когда спора попадает на увлажненную почву, она прорастает, образуя так называемый проросток. Он похож на ветвящуюся нить. На проростке образуются почки, из которых вырастают растения кукушкина льна.
Кукушкин лен — двудомное растение. Это значит, что у него есть мужские и женские растения. На верхушке стеблей мужских растений образуются так называемые антеридии. В них созревают сперматозоиды. На женских растениях образуются архегонии. В каждом архегонии созревает яйцеклетка.
Во время дождей или половодья сперматозоиды подплывают к яйцеклеткам. Происходит оплодотворение и образование зиготы. Вода играет важную роль в жизни мхов. Только благодаря ей возможно половое размножение. Таким образом, в эволюционном развитии мхи не только по строению не очень далеко ушли от водорослей, но и по своему образу жизни.
Из зиготы прямо на материнском растении вырастает ножка, на конце которой находится коробочка с колпачком. У мхов ножка и коробочка с колпачком называются спорофитом, тогда как само зеленое растение называется гаметофитом. На спорофите мхов нет зеленых листьев и, следовательно, он не может сам себя питать. Органические вещества он получает от гаметофита, т. е., можно сказать, паразитирует на нем.
В коробочке спорофита созревают споры. Когда колпачок отпадает, споры рассыпаются. Попав в благоприятные условия, они дают начало новому проростку.
Кукушкин лён — строение, размножение, значение
Кукушкин лен – род листостебельных зеленых мхов отдела моховидные растения. Известно около 100 видов, произрастающих по всей Земле. В виде подушкообразных дерновин кукушкин лен распространен в лесах, тундре, горной местности, на болотах и лугах, зачастую формирует густой полог. Представители данного рода участвуют в заболачивании почв и торфообразовании.
В странах СНГ встречается около 10 видов растений данного рода в северных регионах и средней полосе. Наиболее распространен кукушкин лен обыкновенный, или политрихум обыкновенный. Этими растениями богаты заболачивающиеся леса-долгомошники в тайге, влажные леса и болота.
Строение кукушкина льна. Это многолетний листостебельных мох достаточно крупных размеров с ризоидами в нижней части стебля (примитивными аналогами корней). Первичный горизонтальный стебель развивается без листьев. Вторичный стебель прямостоячий, может быть как простой, так и разветвленный. Длина вторичного стебля составляет, в среднем, 10-15 см, но может достигать 30-40 см. Стебли густо покрыты листьями, каждый из которых на верхней своей поверхности имеет ассимиляционные пластиночки и крупную основную жилку. Нижние листья на стебле развиваются в форме чешуек.
Во внутреннем строении стебля различают примитивную проводящую систему, благодаря чему вода с растворенными питательными веществами перемещается по стеблю. Отдельные удлиненные клетки стебля, в которых отсутствует содержимое, соединены порами аналогично трахеидам высших растений. Их предназначение – транспортировка воды.
Размножение кукушкина льна. Это растение размножается бесполым (побегами, спорами) и половым (гаметами) путем. Гаметофит представляет собой многолетний зеленый побег с листочками и ризоидами (корнеподобными образованиями). Растение образует многочисленные споры. Из каждой споры при благоприятных условиях развивается побег с небольшой продолжительностью жизни, который выглядит как коробочка (спорангий) на ножке. Строение коробочки со спорами отличается от такового других листостебельных мхов. В верхней части она прикрыта колпачком с тонкими волосками и похожа на льняную пряжу. А сами коробочки напоминают кукушку, замершую на шесте. Это определяет название растений данного рода.
Значение кукушкина льна. Благодаря специфическому строению листьев и образованию плотной дернины, распространение приводит к поверхностному накоплению влаги и заболачиванию местности. Также эти мхи участвуют в образовании торфа.
Лабораторная работа№1 Изучение внешнего строения мхов (кукушкин лен, сфагнум)
Дата |
|
| Завуч по УР |
Класс | 7а | 7б | ____________З.И.Яндиева |
Лабораторная работа№1 Изучение внешнего строения мхов (кукушкин лен, сфагнум)
Цель. Ознакомиться с внешним видом зеленого мха и белого мха. Сравнить
строение сфагнума и кукушкина льна. Доказать принадлежность мхов к высшим
споровым растениям.
Ход работы
1. Рассмотрите у зелёного мха (например, кукушкина льна) его стебель с ризоидами, листья, коробочку на ножке.
2. Изучите особенности строения сфагнума — его стебель, листья, коробочку на ножке.
3. Сравните кукушкин лен с мхом – сфагнумом. Отметьте отличия в строении и форме листьев, коробочек, ветвлении стебля.
Сравнение мхов сфагнума и кукушкиного льна (общие черты и отличия сфагнума от кукушкина льна)
Сфагнум | Кукушкин лён | |
Класс | Сфагновые мхи | Листостебельные мхи |
Местообитание | На болотах и в лесу | В лесу и на болотах. На гарях, на вырубках |
Крепление к почве | Корней нет вообще. По мере роста нижняя часть стебля отмирает и оторфовывается | Корней нет, вместо них – ризоиды |
Стебель | Прямой, с веточками | Прямой |
Алгоритм поиска с кукушкой для мультимодальной оптимизации
Интерес к мультимодальной оптимизации быстро растет, поскольку многие практические инженерные задачи требуют локализации нескольких оптимумов в пространстве поиска. С другой стороны, алгоритм поиска с кукушкой (CS) — это простой и эффективный алгоритм глобальной оптимизации, который нельзя напрямую применять для решения задач мультимодальной оптимизации. В этой статье предлагается новый алгоритм мультимодальной оптимизации, называемый мультимодальным поиском с кукушкой (MCS).В рамках MCS исходная CS расширяется за счет мультимодальных возможностей посредством (1) включения механизма памяти для эффективной регистрации потенциальных локальных оптимумов в соответствии с их значением пригодности и расстоянием до других потенциальных решений, (2) модификацией исходного Стратегия индивидуального выбора CS для ускорения процесса обнаружения новых локальных минимумов и (3) включение процедуры очистки для циклического устранения дублированных элементов памяти. Производительность предлагаемого подхода сравнивается с несколькими современными алгоритмами мультимодальной оптимизации с учетом набора тестов из четырнадцати мультимодальных задач.Экспериментальные результаты показывают, что предложенная стратегия способна обеспечить лучшую и даже более стабильную производительность по сравнению с существующими хорошо известными мультимодальными алгоритмами для большинства тестовых задач, при этом избегая любого серьезного ухудшения вычислений.
1. Введение
Оптимизация — это область с приложениями во многих областях науки, техники, экономики и других, где используется математическое моделирование [1]. В общем, цель состоит в том, чтобы найти приемлемое решение целевой функции, определенной в заданном пространстве поиска.Алгоритмы оптимизации обычно делятся на детерминированные и стохастические [2]. Поскольку детерминированные методы обеспечивают только теоретическую гарантию нахождения локального минимума целевой функции, они часто сталкиваются с большими трудностями при решении задач оптимизации [3]. С другой стороны, стохастические методы обычно быстрее определяют глобальный оптимум [4]. Более того, они легко адаптируются к формулировкам «черного ящика» и функциям с крайне плохим поведением, тогда как детерминированные методы обычно основываются, по крайней мере, на некоторых теоретических предположениях о постановке задачи и ее аналитических свойствах (таких как липшицева непрерывность) [5].
Эволюционные алгоритмы (EA), которые считаются членами стохастической группы, были разработаны на основе комбинации правил и случайности, имитирующей несколько природных явлений. К таким явлениям относятся эволюционные процессы, такие как эволюционный алгоритм (EA), предложенный Fogel et al. [6], де Йонг [7] и Коза [8]; генетический алгоритм (ГА), предложенный Холландом [9] и Голдбергом [10]; искусственная иммунная система, предложенная де Кастро и фон Зубеном [11]; и алгоритм дифференциальной эволюции (DE), предложенный Сторном и Прайсом [12].Некоторые другие методы, основанные на физических процессах, включают моделирование отжига, предложенное Киркпатриком и др. [13], подобный электромагнетизму алгоритм, предложенный Бирбилом и Фангом [14], и алгоритм гравитационного поиска, предложенный Рашеди и др. [15]. Кроме того, существуют другие методы, основанные на явлениях поведения животных, такие как алгоритм оптимизации роя частиц (PSO), предложенный Кеннеди и Эберхартом [16], и алгоритм оптимизации колонии муравьев (ACO), предложенный Дориго и др.[17].
Большая часть исследовательской работы по EA направлена на определение глобального оптимума [18]. Несмотря на наилучшие характеристики, глобальный оптимум может быть интегрирован с помощью значений параметров, которые считаются непрактичными или чрезмерно дорогими, что ограничивает их внедрение в реальное приложение. Поэтому с практической точки зрения желательно иметь доступ не только к глобальному оптимуму, но и к как можно большему количеству локальных оптимумов (в идеале — ко всем). В таких обстоятельствах локальный оптимум с приемлемым качеством работы и умеренной стоимостью может быть предпочтительнее дорогостоящего глобального решения с немного лучшей производительностью [19].Процесс поиска глобального оптимума и множественных локальных оптимумов известен как мультимодальная оптимизация.
EA хорошо справляется с поиском одного оптимума, но не может предоставить несколько решений [18]. В схему EA было введено несколько методов для достижения мультимодальной оптимизации, таких как совместное использование пригодности [20–22], детерминированное скопление [23], вероятностное скопление [22, 24], основанное на кластеризации niching [25], процедура клиринга [26] , генетический алгоритм сохранения видов [27] и стратегии элитарной популяции [28].Однако большинство этих методов сопряжено с трудностями, которые необходимо преодолеть, прежде чем их можно будет успешно применять в мультимодальных приложениях. Некоторые выявленные проблемы включают трудности с настройкой некоторых параметров niching, трудности с поддержанием обнаруженных решений в ходе выполнения, дополнительные вычислительные затраты и плохую масштабируемость при высокой размерности. Дополнительная проблема заключается в том, что такие методы разрабатываются для расширения возможностей поиска популярных EA, таких как GA и PSO, которые не могут найти баланс между исследованием и эксплуатацией, в основном для мультимодальных функций [29].Более того, они не исследуют весь регион эффективно и часто страдают от преждевременной конвергенции или потери разнообразия.
В качестве альтернативных подходов другие исследователи использовали искусственные иммунные системы (ИИС) для решения задач мультимодальной оптимизации. Некоторые примеры — алгоритм клонального отбора [30] и искусственная иммунная сеть (AiNet) [31, 32]. Оба подхода используют операторы и структуры, которые пытаются найти множество решений, имитируя поведение естественной иммунной системы.
Каждый эксперт должен решать проблему исследования и использования поискового пространства [33]. Исследование — это процесс посещения совершенно новых точек пространства поиска, в то время как эксплуатация — это процесс уточнения этих точек в окрестностях ранее посещенных мест, чтобы улучшить качество их решения. Чистое исследование снижает точность эволюционного процесса, но увеличивает его способность находить новые потенциальные решения. С другой стороны, чистая эксплуатация позволяет усовершенствовать существующие решения, но отрицательно ведет процесс к локальным оптимальным решениям.
Мультимодальная оптимизация требует достаточного количества исследований агентов популяции в гиперпространстве, чтобы все локальные и глобальные аттракторы могли быть успешно и быстро обнаружены [34, 35]. Однако эффективный алгоритм мультимодальной оптимизации должен демонстрировать не только хорошую тенденцию к исследованию, но и хорошую эксплуатационную способность, особенно на последних этапах поиска, поскольку он должен обеспечивать точную распределенную сходимость к различным оптимумам в ландшафте.Следовательно, способность эксперта находить несколько решений зависит от его способности достичь хорошего баланса между использованием уже найденных элементов и исследованием пространства поиска [36]. До сих пор дилемма разведки и эксплуатации оставалась нерешенной проблемой в рамках EA.
Недавно был предложен новый природный алгоритм, названный алгоритмом поиска с кукушкой (CS) [37], для решения сложных задач оптимизации. Алгоритм CS основан на облигатно-паразитарной стратегии некоторых видов кукушек.Одна из самых мощных функций CS — использование полетов Леви для создания новых возможных решений. При таком подходе возможные решения модифицируются за счет использования множества мелких изменений и иногда больших скачков. В результате CS может существенно улучшить взаимосвязь между разведкой и эксплуатацией, при этом увеличивая свои возможности поиска [38]. Недавние исследования показывают, что CS потенциально намного эффективнее PSO и GA [39]. Такие характеристики мотивировали использование CS для решения различных видов инженерных задач, таких как создание сетки [40], встроенные системы [41], конструкция стального каркаса [42], задачи планирования [43], термодинамика [44] и распределительные сети. [45].
В этой статье представлен новый алгоритм мультимодальной оптимизации, называемый мультимодальным поиском с кукушкой (MCS). Этот метод сочетает в себе алгоритм CS с новым механизмом памяти, который позволяет эффективно регистрировать потенциальные локальные оптимумы в соответствии с их значением пригодности и расстоянием до других потенциальных решений. Первоначальная стратегия отбора CS в основном осуществляется элитарным решением, в котором преобладают лучшие люди. Чтобы ускорить процесс обнаружения потенциальных локальных минимумов в нашем методе, стратегия выбора модифицируется, чтобы на нее влияли индивидуумы, содержащиеся в механизме памяти.В течение каждого поколения в память включаются яйца (особи), которые занимают разные позиции. Поскольку такие люди могут представлять один и тот же локальный оптимум, также включена процедура очистки для циклического устранения дублирующихся элементов памяти. Производительность предлагаемого подхода сравнивается с несколькими современными алгоритмами мультимодальной оптимизации с учетом набора тестов из 14 мультимодальных задач. Экспериментальные результаты показывают, что предложенная стратегия способна обеспечить лучшую и даже более стабильную производительность по сравнению с существующими хорошо известными мультимодальными алгоритмами для большинства тестовых задач, избегая какого-либо серьезного ухудшения вычислений.
Работа организована следующим образом. В разделе 2 объясняется алгоритм поиска с кукушкой (CS), а в разделе 3 представлен предлагаемый подход MCS. Раздел 4 демонстрирует экспериментальный набор и результаты его работы. Наконец, раздел 5 содержит некоторые заключительные замечания.
2. Метод поиска с кукушкой (CS)
CS — один из новейших природных алгоритмов, разработанный Янгом и Деб [37]. CS основан на паразитизме выводков некоторых видов кукушек. Кроме того, этот алгоритм дополнен так называемыми полетами Леви [46], а не простыми изотропными случайными блужданиями.Недавние исследования показывают, что CS потенциально намного эффективнее PSO и GA [39].
Кукушки откладывают яйца в гнезда других птиц-хозяев (обычно других видов) с удивительными способностями, такими как выбор гнезд, содержащих недавно отложенные яйца, и удаление существующих яиц, чтобы увеличить вероятность вылупления их собственных яиц. Некоторые птицы-хозяева способны бороться с этим паразитическим поведением кукушки и выбрасывать обнаруженные инопланетные яйца или строить новое гнездо в другом месте.Эта аналогия с выращиванием кукушек используется для разработки алгоритма CS. Природные системы сложны, и поэтому их нельзя точно смоделировать с помощью компьютерного алгоритма в его базовой форме. Для успешной реализации в компьютерных алгоритмах необходимо упрощение естественных систем. Ян и Деб [39] упростили процесс воспроизводства кукушки тремя идеализированными правилами: (1) Яйцо представляет собой раствор и хранится в гнезде. Искусственная кукушка может отложить только одно яйцо за раз. (2) Кукушка ищет наиболее подходящее гнездо для откладывания яиц (раствор), чтобы максимально увеличить выживаемость своих яиц.Применяется элитарная стратегия отбора, так что только яйца высокого качества (лучшие решения, близкие к оптимальному значению), которые больше похожи на яйца птицы-хозяина, имеют возможность развиваться (следующее поколение) и стать зрелыми кукушками. (3) Количество гнезд хозяев (популяция) фиксируется. Птица-хозяин может обнаружить инопланетное яйцо (худшие решения, отличные от оптимального значения) с вероятностью, и эти яйца будут выброшены или гнездо брошено, и на новом месте построено совершенно новое гнездо.В противном случае яйцо созревает и доживает до следующего поколения. Новые яйца (растворы), отложенные кукушкой, выбирают гнездо Леви полеты вокруг лучших текущих решений.
С точки зрения реализации, в операции CS популяция () яиц (особей) эволюционирует от начальной точки () до общего числа итераций поколения (). Каждое яйцо () представляет собой -мерный вектор, где каждое измерение соответствует переменной решения решаемой задачи оптимизации.Качество каждого яйца (возможное решение) оценивается с помощью целевой функции, конечный результат которой представляет собой значение пригодности. Три разных оператора определяют процесс эволюции CS: (A) полет Леви, (B) замена некоторые гнезда, создавая новые решения, и (C) элитарная стратегия отбора.
2.1. Полет Леви (A)
Одной из самых мощных функций поиска с кукушкой является использование полетов Леви для генерации новых возможных решений (яиц). В соответствии с этим подходом новый вариант решения () создается путем возмущения тока при изменении положения.Чтобы получить случайный шаг, генерируется симметричным распределением Леви. Для производства используется алгоритм Мантеньи [47] следующим образом: где и — -мерные векторы и. Каждый элемент и рассчитывается с учетом следующих нормальных распределений:
Границы | Сходное разнообразие ампликонов генов альфопротеобактерий и нитрогеназ на двух родственных сфагновых мхах
Введение
Северные водно-болотные угодья относятся к старейшим формам растительности с более или менее постоянными условиями на протяжении тысяч лет. Sphagnum — доминирующие торфяники представляют собой один из самых обширных типов северных болот (Дедыш, 2011). Они покрывают 4 миллиона км 2 ок. 3% поверхности Земли и имеют высокую ценность для сохранения биоразнообразия, как резервуар пресной воды, для благосостояния людей и нашего мирового климата из-за своей исключительной роли в связывании углерода (Gorham, 1991; Clymo et al., 1998). Несмотря на свой возраст, эти давно существующие экосистемы чрезвычайно чувствительны к изменению абиотических факторов, связанных с изменением климата (Belyea, Malmer, 2004; Dise, 2009).В качестве доминирующего компонента растительности торфяников мох Sphagnum использовался во всем мире как индикатор изменения климата (Gignac and Vitt, 1994; Whinam and Copson, 2006; Granath et al., 2009). Экологическое значение болот напрямую связано с физическими, морфологическими и химическими характеристиками торфяных мхов Sphagnum ; которые выделяют Sphagnum среди других мхов практически на всех стадиях жизненного цикла (Shaw et al., 2003). Более того, мхи Sphagnum способны изменять среду обитания: живые мхи Sphagna обладают чрезвычайно высокой катионообменной способностью и, следовательно, подкисляют окружающую среду, обменивая связанные с тканью протоны на основные катионы в окружающей воде (Судзиловская и др., 2010). Интересно, что листья Sphagnum очень специализированы: они образуют сеть из живых хлорофилл-содержащих хлорофитов и мертвых гиалоцитов, не содержащих клеточного содержимого, которые ответственны за их высокую водоудерживающую способность. Sphagnum видов также продуцируют видоспецифичные вторичные биологически активные метаболиты, влияющие на колонизацию микробов (Opelt et al., 2007a).
Мхи Sphagnum заселены разнообразными бактериальными сообществами. Популяции микробов, участвующие в циклическом цикле CH 4 , i.е., метанотрофные бактерии (Dedysh et al., 1998; Dedysh, 2002; Raghoebarsing et al., 2005; Larmola et al., 2010; rev. Dedysh, 2011), а также метаногены, включая археи (Horn et al., 2003; Freitag et al., 2010) привлекли внимание исследователей в связи с их важной функцией для выбросов метана. Недавно мы смогли показать, что живые мхи Sphagnum в большом количестве колонизированы определенными микроорганизмами, которые выполняют другие важные функции, такие как снабжение питательными веществами и защита от патогенов для роста и здоровья мха (Opelt et al., 2007а, б). Было обнаружено чрезвычайно сильное влияние вида Sphagnum на структуру микробного разнообразия, и это разнообразие передается непосредственно от спорофита (внутри капсулы спорангия) к гаметофиту и , наоборот, (Брагина и др., 2011). ). В последнем мы проанализировали бактериальные сообщества двух вездесущих видов Sphagnum , S. magellanicum и Sphagnum fallax на трех альпийских болотах в Австрии. Чрезвычайно высокие различия между бактериальными сообществами обоих Sphagna были обнаружены с помощью комбинации методов независимо от места.Например, был идентифицирован различительный спектр бактерий: в то время как альфа- и гамма-протеобактерии доминировали в S. magellanicum , S. fallax в основном колонизировали Verrucomicrobia, Planctomycetes и Alphaproteobacteria. Кроме того, бактериальные сообщества в значительной степени определялись абиотическими факторами (богатство питательными веществами и pH) и сильно коррелировали с составом сообществ высших растений. Специфическое микробное разнообразие, связанное с очень разнообразным родом Sphagnum (Daniels and Eddy, 1985), в значительной степени неизвестно, но важно для понимания и защиты Sphagnum в болотных экосистемах.
Целью данной работы было изучение структуры и функции бактерий, ассоциированных с Sphagnum , с целью понять масштабы и степень специфичности хозяина. Таким образом, были отобраны два филогенетически близких и широко распространенных вида с перекрывающимися микронишами, но различной трофической специализацией: Sphagnum angustifolium (Warnst.) C. E. O. Jensen и S. fallax H. Klinggr. (Дэниелс и Эдди, 1985; Флатберг, 1992; Састад и др., 1999).Оба вида Sphagnum впервые были охарактеризованы по профилю вторичных метаболитов. Для изучения бактериальных сообществ был применен полифазный подход с особым вниманием к Alphaproteobacteria и азотфиксирующим бактериям: (i) микробные отпечатки пальцев с помощью ПЦР-однонитевого конформационного полиморфизма (SSCP) с применением универсальных и группо-специфичных праймеров, направленных на ген 16S рРНК. и специфичные для гена праймеры нитрогеназы ( nifH ), (ii) флуоресцентная гибридизация in situ с универсальными и группоспецифичными зондами в сочетании с флуоресценцией in situ гибридизация конфокальная лазерная сканирующая микроскопия (FISH – CLSM) и анализ изображений, ( iii) глубокое секвенирование Alphaproteobacteria и (iv) функциональный подход к анализу потенциала фиксации азота генами нитрогеназы ( nifH ) в библиотеках ампликонов.
Материалы и методы
Процедура отбора проб
Взрослые гаметофиты S. angustifolium (секция Cuspidata ) и S. fallax (секция Cuspidata ) были взяты из болота Pürgschachen Moor (Лизен, 1,7 км к юго-западу от Арднинга, N47 ° 34,789′E14 °. 2017 ′) в Австрии в июле и ноябре 2010 года. В этом болоте S. angustifolium имеет более широкую экологическую амплитуду и растет в мезотрофных влажных лощинах с некоторым влиянием грунтовых вод, а также в более омбротрофных торосах над уровнем грунтовых вод. S. fallax растут только во влажных мезотрофных условиях, в которых мы собрали образцы обоих видов в сопоставимых экологических ситуациях. PH окружающей торфяной воды был измерен во всех точках отбора проб и показал средние значения 4,00 (SD, 0,15) для S. fallax и 4,04 (SD, 0,31) для S. angustifolium . Всего было собрано восемь независимых повторностей на видов Sphagnum , состоящих из составных выборок из 15-20 проростков, и они хранились отдельно.Приблизительная длина отобранных растений составляла 14,5 см (SD, 2,3 см) для S. fallax и 15,1 см (SD, 2,5 см) для S. angustifolium . Восемь точек отбора проб располагались на расстоянии не менее 10 м, в то время как оба вида находились на расстоянии не более 1 м в каждой отдельной точке. Живые зеленые части проростков помещали в стерильные полиэтиленовые пакеты и транспортировали охлажденными в лабораторию.
Химический анализ сфагнума Вторичных метаболитов
Спектры вторичных метаболитов образцов Sphagnum были проанализированы с использованием высокоэффективной жидкостной хроматографии с УФ-детектированием фотодиодной матрицы (HPLC-PDA), как описано ранее (Opelt et al., 2007а).
Выделение тотальной ДНК сообщества
Бактериальная фракция, ассоциированная с гаметофитами, была извлечена в соответствии с модифицированным протоколом Opelt and Berg (2004). Вкратце, 5 г растительного материала физически разрушали стерильным пестиком и ступкой и повторно суспендировали в 10 мл 0,85% NaCl. Два миллилитра суспензии центрифугировали при 13000 об / мин в течение 20 минут при 4 ° C, и осадок использовали для выделения ДНК всего сообщества. Для механического лизиса клетки дважды гомогенизировали в FastPrep ® FP120 Instrument (MP Biomedicals) в течение 30 с на скорости 5.0. Полученную ДНК очищали с использованием набора FastDNA ® SPIN для почвы (MP Biomedicals) в соответствии с протоколом производителя. Окончательные аликвоты ДНК всего сообщества были далее применены в подходах на основе ПЦР.
Микробный отпечаток пальца с помощью ПЦР-SSCP
Отпечатки ассоциированных со мхом бактериальных сообществ были выполнены с помощью SSCP на основе ПЦР, описанного Schwieger and Tebbe (1998). Гены 16S рРНК Бактерии амплифицировали с универсальными бактериальными праймерами Com1 / Unibac-II-927r P (Schwieger, Tebbe, 1998; Zachow et al., 2008). Набор специфичных для Alphaproteobacteria праймеров ADF681F / 1492r, за которым следует ADF681F / 927r P , применяли с использованием полугнездового протокола (Blackwood et al., 2005). Фрагменты гена бактериальной нитрогеназы ( nifH ) амплифицировали во вложенном подходе с праймерами nifh4 / 19F, nifh21 / nifh32 P (Yeager et al., 2004). Ампликоны разделяли с использованием системы TGGE Maxi (Biometra) при 400 В и 26 ° C в акриламидном геле с последующим окрашиванием серебром.
Конформационный полиморфизм цепи основан на различиях в конформации одноцепочечных фрагментов ДНК.Электрофоретическая подвижность одноцепочечных фрагментов ДНК зависит от их трехмерной конформации. Каждый из продуктов амплификации идентифицировали по его электрофоретическому расстоянию на геле SSCP и количеству фрагментов ДНК. В зависимости от расстояния между полосами гели SSCP виртуально были разделены на операционные таксономические единицы (OTU). Оценивали наличие или отсутствие отдельных полос амплифицированного продукта ДНК в каждой группе. Полученная матрица использовалась для статистического сравнения (см. Статистику).
Флуоресцентный in situ Гибридизация и конфокальная лазерная сканирующая микроскопия
Одиночные гаметофиты S. angustifolium и S. fallax фиксировали 4% забуференной солью параформальдегид / фосфат (3: 1, об. / Об.). Отделенные листья окрашивали методом FISH в пробирке (Grube et al., 2009). Флуоресцентно меченый зонд для нацеливания рРНК ALF968, специфичный для Alphaproteobacteria (Loy et al., 2007), применяли в комбинации с эквимолярной смесью универсальных бактериальных зондов EUB338, EUB338II, EUB338III (Amann et al., 1990; Daims et al., 1999). Образцы Sphagnum были последовательно гибридизированы с ALF968 (41 ° C, 45% формамид), а затем с EUB338 / EUB338II / EUB338III (41 ° C, 15% формамид). Отрицательный контроль гибридизовали с нецелевым зондом NON-EUB (Amann et al., 1990) в тех же жестких условиях, что и для положительных зондов FISH.
Конфокальная лазерная сканирующая микроскопия выполнялась с использованием конфокального микроскопа Leica TCS SPE (Leica Microsystems). Флуоресцентные красители Cy3 и Cy5, меченные для зондов FISH, последовательно возбуждали лазерными лучами с длиной волны 532 и 635 нм соответственно; излучаемый свет регистрировался в диапазоне 556–607 и 657–709 нм соответственно.Дополнительный канал (возбуждение 488 нм; диапазон излучения 508–556 нм) использовали для регистрации автофлуоресценции клеток мха. Усиление и смещение фотоумножителя были индивидуально оптимизированы для каждого канала и каждого поля обзора, чтобы улучшить соотношение сигнал / шум. Конфокальные стопки были получены с помощью объектива Leica ACS APO 40X OIL CS (NA: 1,15) и объектива Leica ACS APO 63X OIL CS (NA: 1,30) с шагом Z 0,4–0,8 мкм. Трехмерные реконструкции были созданы с помощью программного обеспечения Imaris 7.0 (битовая плоскость).
Глубокое секвенирование и биоинформатический анализ
Разнообразие Alphaproteobacteria и азотфиксирующих бактерий, ассоциированных с видами Sphagnum , было глубоко изучено с помощью метода пиросеквенирования со штрих-кодом. ДНК всего сообщества амплифицировали с набором специфичных для Alphaproteobacteria праймеров ADF681F / Unibac-II-927r (Blackwood et al., 2005; Zachow et al., 2008) и nifH, ген-специфичных праймеров nifh4 / nifh5, nifh2. / nifh3 во вложенном подходе (Zehr and Turner, 2001) с использованием Taq- & Go ™ Ready-to-use PCR Mix (MP Biomedicals).Дублированные продукты ПЦР из всех шаблонов очищали с помощью геля Wizard ® SV и системы очистки PCR (Promega). Ампликоны каждого Sphagnum sp. были объединены и подвергнуты пиросеквенированию с использованием платформы Roche / 454 GS FLX + Titanium, выполненной GATC Biotech (Констанц, Германия).
Библиотеки ампликонов гена 16S рРНК, специфичные для Alphaproteobacteria, анализировали, как указано Bragina et al. (2011). Вскоре исходные данные секвенирования были отфильтрованы по качеству и длине (≥150 п.н.).Анализ разреженности был проведен для кластеров филотипов 97, 95 и 90% сходства с использованием инструментов RDP’s Pyrosequencing Pipeline (Cole et al., 2009). Наборы данных были нормализованы до одинакового количества последовательностей. Оценки богатства и индексы разнообразия рассчитывались в пакете программного обеспечения с открытым исходным кодом QIIME (Caporaso et al., 2010). Классификация прочтений проводилась с использованием конвейера BLAT в веб-интерфейсе SnoWMAn версии 1.11 с 80% доверительным порогом.
Библиотеки Ampliconгена нитрогеназы ( nifH ) были исследованы с использованием FunGene Pipeline сервера RDP с параметрами, указанными Farnelid et al.(2011). Последовательности праймеров были обрезаны, и считывания низкого качества и более короткие 200 п.н. были удалены. Отфильтрованные чтения были переведены в аминокислотные последовательности и обрезаны до 60 аминокислотных остатков. Дальнейшие анализы проводились на аминокислотных последовательностях. Для разрежения наборы данных были сгруппированы с порогами сходства 100, 96 и 92%. Оценки богатства и индексы разнообразия были рассчитаны для подмножеств, нормализованных к одинаковому количеству последовательностей с помощью программного обеспечения QIIME. Композиционное разнообразие сравнивали по индексам Соренсена ( C s ) и Шеннона ( H ′) на уровне сходства 96%.Филогенетический анализ был выполнен для кластеров 92% сходства с ≥10 последовательностями. Эталонные последовательности были получены с использованием алгоритма NCBI TBLASTN, а филогенетическое дерево было построено, как описано ранее (Брагина и др., 2011).
Статистика
Компьютерный анализ профилей SSCP был выполнен с использованием программного пакета GelCompare II версии 5.1 (Applied Maths). Матрицы сходства были построены на основе коэффициентов корреляции Пирсона (–), а кластерный анализ был выполнен методом невзвешенных парных групп со средними связями (UPGMA).Профили SSCP микробных сообществ, созданные с помощью универсальных бактериальных праймеров и праймеров, специфичных для гена nifH , были дополнительно использованы для многомерного анализа. Отдельные полосы ДНК, характеризующиеся относительным положением и численностью на геле, были определены как OTU и использовались в качестве переменных отклика для анализа соответствия с исключенным трендом (DCA) Canoco 4.5 для Windows (Lepš and Smilauer, 2003).
Результаты
Профилирование сфагнума Вторичных метаболитов
Высокоэффективная жидкостная хроматография-PDA профилирование растительных экстрактов дало заметно похожие хроматограммы и спектры преобладающих пиков вторичных метаболитов для обоих видов Sphagnum .УФ-спектры предполагают наличие различных фенолов и производных индола, включая сфагновую кислоту (пик 7; рис. 1). Единственным заметным отличием было более выраженное накопление конъюгатов флавоноидов (нарингенина и апигенина) у S. angustifolium . Позднее время удерживания препятствовало гликозидам, но более липофильным молекулам. Результат свидетельствовал об общей химической основе для установления сходного микробного разнообразия сравниваемых видов мхов.
Рисунок 1.Высокоэффективная жидкостная хроматография — УФ-профили вторичных метаболитов S. angustifolium и S. fallax при 229 нм (УФ-спектры (200–400 нм; 1, фенол; 2, 5, 6, производное индола; 3, 4) — фенолы; 7, 9 — сфагновая кислота; 8 — фенол; 10 — флавоноиды нарингенинового ряда; 11, 12 — флавоноиды апигенина .
Молекулярный фингерпринтинг микробных сообществ
В подходе молекулярного фингерпринтинга мы использовали определенный набор праймеров: универсальные праймеры, чтобы получить обзор всего бактериального сообщества, связанного с Sphagnum , праймеры для Alphaproteobacteria, потому что это повсеместный и космополитический филогенетический класс, связанный с Sphagnum (Bragina и другие., 2011) и праймеров nifH для определения азотфиксирующих бактерий как важной функциональной группы (Opelt et al., 2007a). Все отпечатки пальцев SSCP, полученные с помощью универсальных и группоспецифичных праймеров, выявили высокое сходство микробных сообществ, связанных с S. fallax и S. angustifolium (рис. 2). Анализ профилей бактерий UPGMA показал минимальный уровень сходства в 95,5% сообществ, связанных с мохообразными. Паттерны альфапротеобактерий сформировали общий кластер на 87.Сходство 1%. Отпечатки гена nifH сгруппированы в две группы со сходством 41,2%. Внутри каждого кластера образцы обоих Sphagna сгруппированы вместе. Это совпадение микробных сообществ было подтверждено статистически с помощью анализа соответствия без тренда (Рисунок 3).
Рисунок 2. Метод невзвешенных парных групп со средними дендрограммами связей бактериальных сообществ, ассоциированных с S. angustifolium (SA, квадраты) и S.fallax (SF, кружочки) . Дендрограммы были созданы из профилей PCR-SSCP бактерий (A) , Alphaproteobacteria (B) и бактериальных генов nifH (C) с использованием метода невзвешенных парных групп со средними связями (UPGMA). Цифры в круглых скобках обозначают дубликаты. Двуглавые вертикальные стрелки указывают на сходство группировок.
Рис. 3. Анализ соответствия без тренда (DCA, косвенный одномодальный градиентный анализ) операционных таксономических единиц (OTU), идентифицированных по отпечаткам пальцев сообщества SSCP .Собственные значения первой и второй оси равны 0,244 и 0,141 соответственно; сумма всех собственных значений 1.715. Черные звезды и ромбы показывают расположение 16 образцов, цветные кружки — расположение 111 OTU на графике. Цвета указывают на его предпочтение к Sphagnum angustifolium (ang) или S. fallax соответственно.
Модели колонизации микробов
Sphagnum гаметофитов, характеризующихся уникальной морфологией, были изучены на предмет микробной колонизации с помощью FISH.Опять же, мы применили универсальные и специфичные для Alphaproteobacteria пробы по указанным выше причинам. Стеблевые и ветвящиеся листья обычно различаются по размеру и форме, но в равной степени они образованы диморфными клетками листа, в которых большие пустые гиалиновые клетки, перфорированные порами, заключены в сеть более узких хлорофиллозных клеток (рис. 4A, D). Наблюдение с помощью CLSM обоих типов листьев показало плотную колонизацию бактериальными колониями внутреннего пространства гаметофитов, как показано для ветвей листьев на фиг. 4B, E.Клетки альфапротеобактерий представляли до 50% обнаруженных бактериальных колоний. Трехмерная реконструкция полученных изображений подтвердила, что бактерии в основном занимали мертвые гиалиновые клетки. Рисунки 4C, F. В заключение, подход FISH – CLSM показал сходные паттерны колонизации для S. angustifolium и S. fallax бактериальными сообществами.
Рисунок 4. Локализация бактерий в гаметофитах мха .Флуоресцентный in situ гибридизация S. angustifolium (A – C) и S. fallax (D – F) листьев показала колонизацию гиалиновых клеток Alphaproteobacteria. Изображения, полученные с помощью конфокальной лазерной сканирующей микроскопии (CLSM) (A, B, D, E) : фиолетовый — клеточные стенки клеток Sphagnum ; зеленый — хлорофиллсодержащие хлороцитов Sphagnum ; желтый — Alphaproteobacteria; красный — другие бактерии. 3D компьютерные реконструкции изображений CLSM с использованием Imaris7.0 (C, F) : синий — моховая ткань; красный: Alphaproteobacteria; зеленый: другие бактерии. Масштабная линейка = 20 мкм.
Глубокое секвенирование Alphaproteobacteria и NIFH Гены
Alphaproteobacteria и азотфиксирующие бактерии были отобраны для получения более глубокого понимания с помощью подхода пиросеквенирования. Библиотеки ампликонов гена 16S рРНК, специфичные для Alphaproteobacteria, были разрежены, как показано на рисунке 5. Оценка насыщенности нормализованных наборов данных показала, что усилия по пиросеквенированию достигли 66.0–74,2% от оценочного богатства для кластеров с 90% сходством (Таблица 1). Кластеры с 95-процентным и 97-процентным сходством отражали 49,3–51,2 и 45,4–46,2% предполагаемого богатства, соответственно.
Рис. 5. Кривые разрежения для библиотек ампликонов Sphagnum образцов . Кривые насыщения представлены для образцов S. angustifolium (SA) и S. fallax (SF). Alphaproteobacteria (A) Наборы данных и nifH (B) были сгруппированы с определенными границами сходства.
Таблица 1 . Оценки богатства и индексы разнообразия для библиотек ампликонов образцов Sphagnum a .
Таксономический состав популяций альфа-протеобактерий по сравнению с рангами семейств и родов был в значительной степени сходным для Sphagnum spp. (Рисунок 6). Доминирующее семейство Acetobacteraceae преобладали родами Acidocella , Acidisphaera и Rhodopila .Внутри семейств Sphingomonadaceae и Rhodospirillaceae большинство членов принадлежало к видам Novosphingobium spp. и Magnetospirillum spp. соответственно. Вместе с тем, состав и соотношение субдоминанта Caulobacteraceae варьировалось между мхами. Семейство было более многочисленным в образце S. fallax и состояло из рода Phenylobacterium (обнаружено повсюду) и рода Caulobacter (уникального для S.fallax ). Разнообразие видов оценивалось по индексу разнообразия Шеннона ( H ′) для кластеров с 97% сходством. Сравнение значений индекса выявило несколько большее разнообразие Alphaproteobacteria для S. fallax (4.60), чем для S. angustifolium (4.18).
Рис. 6. Таксономическая классификация Alphaproteobacteria, связанных с S. angustifolium и S. fallax .Считывания пиросеквенирования классифицируются по семейству (A) и уровню рода (B) с порогом достоверности 80%. Группы менее 1% относительной численности включены в Прочие . Разноцветные диаграммы у легенды показаны для каждого образца соответственно.
Согласно базе данных NCBI, идентифицированные роды альфа-протеобактерий включают бактерии, известные своей азотфиксацией. В частности, в обеих библиотеках были обнаружены роды Bradyrhizobium , Acetobacter и Beijerinckia , в то время как в S.Fallax библиотека.
Анализ разреженности библиотек генов нитрогеназы привел к аналогичным профилям насыщения образцов Sphagnum (рис. 5). Нормализованные наборы данных представляли 61,0–62,7% от оценочного богатства при 92% сходстве (таблица 1). Кластеры с 96% -ным сходством охватывают 55,2–66,6%, а уникальные кластеры отражают 19,8–22,1% оценочного богатства, соответственно. Композиционное разнообразие оценивалось с применением ограничения сходства 96% для ограничения кластеров.Индекс сходства Соренсена показал, что образцы имели идентичность на 53%. Разнообразие нитрогеназ, оцененное Шенноном, снова было немного выше для S. fallax (7,59), чем для S. angustifolium (7,20; Таблица 1).
Филогенетический анализ состава NifH показал, что извлеченные последовательности длиной 60 аминокислот были распределены между каноническими кластерами I, III и подкластером IA (рис. 7). В наиболее многочисленном кластере I, включающем последовательности Proteobacteria, и Cyanobacteria, преобладали Alphaproteobacteria.Согласно анализу BLAST, наиболее распространенные альфа-протеобактериальные ампликоны на ≥95% сходны с видами Bradyrhizobium , Azorhizobium , Rhizobium , Methylobacterium , Rhodocista и Acetobacter . Значительная часть ампликонов продемонстрировала 96–100% идентичность с эталонными последовательностями Methylocella , Methylocapsa и Beijerinckia . Обнаруженные бетапротеобактерии преобладали среди Burkholderia spp.(≥95% сходства). Незначительная часть ампликонов кластера I была связана с цианобактериями родов Anabaena и Tolypotrix (100% сходство). Подкластер IA содержал последовательности, на 96–100% сходные с Geobacter sp. Внутри кластера III ампликоны сгруппированы по ссылочным видам Spirochaeta и Thermincola (сходство 82–93%). В заключение, библиотеки ампликонов nifH из Sphagnum -ассоциированных микробных сообществ характеризовались высоким разнообразием и изобилием ампликонов альфа-протеобактерий.
Фигура 7. Филогенетический состав библиотек ампликонов гена нитрогеназы ( nifH ) S. angustifolium (SA, кружки) и S. fallax (SF, квадраты) . Соседнее филогенетическое дерево было построено как с одной репрезентативной последовательностью для кластера NifH (92% сходства), так и с ближайшими ссылочными последовательностями (номера доступа в скобках). Частичная последовательность светонезависимой субъединицы L фотохлорофиллидредуктазы (BchL) из Chlorobaculum tepidum (Acc.№ AAG12203) использовалась как чужая группа. Надежность топологии дерева оценивалась 100 повторными попытками начальной загрузки (значения начальной загрузки не показаны). Обилие и филогенетическая принадлежность гроздей указаны в легенде. Масштабная линейка = 0,1 замены на сайт.
Обсуждение
Мхи Sphagnum образуют выдающуюся группу из Bryophyta ; они уникальны по своим морфологическим особенностям и особенностям развития на каждой стадии жизненного цикла (Shaw et al., 2003). Физико-химические характеристики мха Sphagnum больше, чем для других растений, связаны с экологией и функцией северных торфяников. В этом исследовании мы показали такую же глубокую связь между проростками мха и связанными с ними бактериальными сообществами. В многостороннем подходе, примененном к изучению структуры и функции бактерий, между S были обнаружены лишь незначительные, но не статистически значимые различия. angustifolium и S.fallax , два торфяных мха, которые имели сходные экологические условия внутри экосистемы болота.
Это высокое сходство контрастирует с предыдущими исследованиями бактерий, ассоциированных с Sphagnum , различных экологических амплитуд (Opelt et al., 2007c; Bragina et al., 2011). Основное различие между предыдущими исследованиями и настоящим заключается в близком таксономическом и экологическом родстве исследованных S. angustifolium и S. fallax в отличие от S.magellanicum . S. magellanicum принадлежит к другой секции в пределах рода Sphagnum (секция Sphagnum ) и типична для сильнокислых, олиготрофных и омбротрофных местообитаний, тогда как S. angustifolium и S. fallax (секция Cuspidata ) растут в слабокислых, более мезотрофных условиях под влиянием минеральных грунтовых вод (Daniels and Eddy, 1985). Также с морфологической точки зрения S. angustifolium и S.fallax трудно различить, и раньше оба таксона считались разновидностями одного вида. S. recurvum P. Beauv (Smith, 1978). В отличие от S. magellanicum (Opelt et al., 2007a), S. angustifolium и S. fallax характеризуются сходными вторичными метаболитами. Совпадение общих свойств мхов было обнаружено и для бактериального сообщества. Рисунок 3 показывает не только высокую степень сходства между микробными сообществами, но и дифференциацию более S.fallax (отрицательно по первой оси) и еще S. angustifolium (положительно по первой оси), предпочитая бактерии. Таким образом, ситуацию, показанную в качестве примера на рисунке 3, можно интерпретировать как раннее состояние спецификации бактериальных сообществ, соответствующее раннему состоянию дифференциации видов хозяев. Интересно, что наши результаты также объясняют различия между теорией видоспецифичных сообществ Sphagnum , установленной Opelt et al. (2007c) и Брагина и др.(2011) и результаты, полученные Larmola et al. (2010), которые выявили только абиотические факторы. Для метанотрофов они обнаружили для трансплантированных Sphagnum бактериальный паттерн и активность, типичные для абиотических параметров целевого участка. Однако это был искусственный эксперимент; большинство ок. 300 Sphagnum видов имеют очень узкие экологические амплитуды и не могут расти в природе в других условиях (Daniels and Eddy, 1985). В северных водно-болотных угодьях, которые относятся к старому типу растительности с более или менее экстремальными, но постоянными условиями в течение тысяч лет, мхи Sphagnum установили высокоспецифический и адаптированный симбиоз со связанными с ними микробами.
Каковы экологические последствия этой специфики микробного сообщества? Хорошо адаптированные к абиотическим параметрам места своего обитания, мхи Sphagnum вместе со своим микробиомом в качестве так называемых «метаорганизмов» выполняют важные функции для экосистемных услуг. Эти функции могут выполняться только в сотрудничестве с соответствующим микробным сообществом. Последний отвечает за фиксацию азота для растения-хозяина, за растворение люминофора, а также за получение углерода из метана, доставляемого торфом (Raghoebarsing et al., 2005; Opelt et al., 2007a). Исследования показали, что окисление CH 4 метанотрофными микробами, находящимися в слое Sphagnum , контролируется факторами окружающей среды, то есть уровнем грунтовых вод и температурой (Larmola et al., 2010), но помимо этого скорость была специфичной для Sphagnum. вида (Gifford et al., 2011). Взятые вместе, специфика микробного сообщества имеет важное значение для жизни в условиях экстремальных и сильно меняющихся экологических градиентов внутри экосистемы болота и для выполнения экологических функций.Болотная экосистема сложнее, чем считалось ранее, но это важно знать для поддержания болотных экосистем на северных водно-болотных угодьях. Высокая специфичность, узкая экологическая амплитуда и тесная взаимосвязь могут быть одной из причин того, что Sphagnum очень чувствителен к изменению биотических параметров, связанных с изменением климата.
Мало что известно об особой экологии бактерий, ассоциированных с Sphagnum , которые в значительной степени еще не культивируются (Дедыш, 2011).Доминирующие таксоны альфа-протеобактерий, ассоциированные со Sphagna, известны как ацидофильные или ацидотолерантные бактерии, способные расти химио-органотрофно или фототрофно и выживать в олиготрофных условиях. Sphagnum -ассоциированные микробные сообщества должны быть включены в повестку дня по сохранению биоразнообразия и использоваться для прогнозирования микробной экологии в соответствии с требованиями Bodelier (2011).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Это исследование финансировалось Австрийским научным фондом FWF за счет гранта Габриэле Берг.
Сноски
Список литературы
Аманн Р. И., Биндер Б. Дж., Олсон Р. Дж., Чисхолм С. В., Деверо Р. и Шталь Д. А. (1990). Комбинация олигонуклеотидных зондов, нацеленных на 16S рРНК, с проточной цитометрией для анализа смешанных микробных популяций. Заявл. Environ. Microbiol. 56, 1919–1925.
Pubmed Реферат | Pubmed Полный текст
Белеа, Л.Р., Малмер Н. (2004). Связывание углерода в торфяниках: закономерности и механизмы реагирования на изменение климата. Glob. Чанг. Биол. 10, 1043–1052.
CrossRef Полный текст
Боделье, П. Л. (2011). На пути к пониманию, управлению и защите микробных экосистем. Фронт. Microbiol. 2:80. DOI: 10.3389 / fmcib.2011.00080
CrossRef Полный текст
Брагина А., Берг К., Кардинале М., Щербаков А., Чеботарь В., Берг Г. (2011). Sphagnum Мхи обладают высокоспецифическим бактериальным разнообразием на протяжении всего своего жизненного цикла. ISME J . DOI: 10.1038 / ismej.2011.151
CrossRef Полный текст
Капорасо, Дж. Г., Кучински, Дж., Стомбо, Дж., Биттингер, К., Бушман, Ф. Д., Костелло, Э. К., Фирер, А., Пена, А. Г., Гудрич, Дж. К., Гордон, Дж., Хаттли, Джорджия, Келли , ST, Найтс, Д., Кениг, Дж. Э., Лей, Р. Э., Лозупоне, Калифорния, Макдональд, Д., Muegge, BD, Pirrung, M., Reeder, J., Sevinsky, JR, Tumbaugh, PJ, Walters, WA, Widmann, J., Yatsunenko, T., Zaneveld, J., and Knight, R. (2010) . QIIME позволяет анализировать высокопроизводительные данные секвенирования сообщества. Nat. Методы 7, 335–336.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Климо, Р. С., Турунен, Дж., И Толонен, К. (1998). Накопление углерода в торфяниках. Oikos 81, 368–388.
CrossRef Полный текст
Коул, Дж.Р., Ван, К., Карденас, Э., Фиш, Дж., Чай, Б., Фаррис, Р.Дж., Кулам-Сайед-Мохидин, А.С., МакГаррелл, Д.М., Марш, Т., Гаррити, GM, и Тидже , JM (2009). Проект базы данных рибосом: улучшенное выравнивание и новые инструменты для анализа рРНК. Nucleic Acids Res. 37, D141 – D145.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Даймс, Х., Брюль, А., Аманн, Р., Шлейфер, К. Х., и Вагнер, М. (1999). Доменно-специфического зонда EUB338 недостаточно для обнаружения всех бактерий: разработки и оценки более полного набора зондов. Syst. Appl. Microbiol. 22, 434–444.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Дэниэлс Р. Э. и Эдди А. (1985). Справочник Europaen Sphagna . Аберистуит: Кембрийские новости.
Дедыш С. Н. (2011). Выращивание некультивируемых бактерий из северных водно-болотных угодий: полученные знания и оставшиеся пробелы. Фронт. Microbiol. 2: 184. DOI: 10.3389 / fmcib.2011.00184
Pubmed Реферат | Pubmed Полный текст
Дедыш, С.Н., Паников, Н. С., Тидже, Дж. М. (1998). Ацидофильные метанотрофные сообщества торфяников Sphagnum . Заявл. Environ. Microbiol. 64, 922–929.
Pubmed Реферат | Pubmed Полный текст
Фарнелид, Х., Андерссон, А. Ф., Бертилссон, С., Аль-Суд, В. А., Хансен, Л. Х., Соренсен, С., Стюард, Г. Х., Хэгстром, А., и Риман, Л. (2011). В ампликонах генов нитрогеназы из глобальных морских поверхностных вод преобладают гены нецианобактерий. PLoS ONE 6, e19223. DOI: 10.1371 / journal.pone.0019223
CrossRef Полный текст
Флатберг, К. И. (1992). Европейские таксоны в комплексе Sphagnum recurvum . 2. Измененные описания Sphagnum brevifolium и S. fallax . Lindbergia 17, 96–110.
Фрейтаг, Т. Э., Тоет, С., Инесон, П., и Проссер, Дж. И. (2010). Связь между потоком метана и транскрипционной активностью метаногенов и окислителей метана в сплошном торфяном болоте. FEMS Microbiol. Ecol. 73, 157–165.
Pubmed Реферат | Pubmed Полный текст
Гиффорд Дж., Хупес М., Лармола Т., Варнер Р., Бубье Дж. И Янг Б. (2011). Роль сфагновых мхов в окислении метана в болоте умеренного пояса . Диссертация, Колледж Маунт-Холиок, Южный Хэдли, Массачусетс.
Жиньяк, Л. Д., и Витт, Д. Х. (1994). Ответные меры северных торфяников на изменение климата: влияние на мохообразные. J. Hattori Bot.Лаборатория. 75, 119–132.
Горхэм, Э. (1991). Северные торфяники: роль в углеродном цикле и вероятные реакции на потепление климата. Ecol. Appl. 1, 182–195.
CrossRef Полный текст
Grube, M., Cardinale, M., Vieira de Castro, J., Müller, H., and Berg, G. (2009). Видовое структурное и функциональное разнообразие бактериальных сообществ при симбиозах лишайников. ISME J. 3, 1105–1115.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Хорн, М.А., Маттис, К., Кюзель, К., Шрамм, А., и Дрейк, Х. Л. (2003). Гидрогенотрофный метаногенез умеренно кислотоустойчивыми метаногенами кислого торфа с выделением метана. Заявл. Environ. Microbiol. 69, 74–83.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Лармола, Т., Туиттила, Э. С., Тиирола, М., Нюкянен, Х., Мартикайнен, П. Дж., Юрьяля, К., Туомивирта, Т., и Фритце, Х. (2010). Роль мхов Sphagnum в круговороте метана северных болот. Экология 91, 2356–2365.
Лепш, J., и Смилауэр, П. (2003). Многомерный анализ экологических данных с использованием Canoco . Кембридж: Издательство Кембриджского университета.
Опельт, К., и Берг, Г. (2004). Разнообразие и антагонистический потенциал бактерий, связанных с мохообразными из бедных питательными веществами местообитаний побережья Балтийского моря. Заявл. Environ. Microbiol. 70, 6569–6579.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Опельт, К., Чобот, В., Хадачек, Ф., Шёнманн, С., Эберл, Л., и Берг, Г. (2007a). Исследования структуры и функции бактериальных сообществ, ассоциированных с мхами Sphagnum . Environ. Microbiol. 91, 2795–2809.
CrossRef Полный текст
Опельт К., Берг К. и Берг Г. (2007b). Род мохообразных Sphagnum является резервуаром мощных и необычных антагонистов и потенциально факультативных патогенов человека. FEMS Microbiol.Ecol. 61, 38–53.
CrossRef Полный текст
Опельт К., Берг К., Шёнманн С., Эберл Л. и Берг Г. (2007c). Высокая специфичность, но контрастное биоразнообразие Sphagnum -ассоциированных бактериальных и растительных сообществ в болотных экосистемах независимо от географического региона. ISME J. 1, 502–516.
CrossRef Полный текст
Raghoebarsing, A. A., Smolders, A. J. P., Schmid, M. C., Rijpstra, W. I. C., Wolters-Arts, Derksen, J. M., Jetten, M. S. M., Schouten, S., Damsté, J. S. S., Lamers, L.P.M., Roelofs, J. G. M., Op den Camp, H. J. M., and Strous, M. (2005). Метанотрофные симбионты обеспечивают углерод для фотосинтеза в торфяных болотах. Природа 436, 1153–1156.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Састад, С. М., Бард, П., и Кьетил, Д. (1999). Генетические эффекты, специфичные для среды обитания, на скорость роста и морфологию в зависимости от градиента pH и уровня воды в популяции мха Sphagnum angustifolium (Sphagnaceae). Am. J. Bot. 86, 1687–1698.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Schwieger, F. и Tebbe, C.C. (1998). Новый подход к использованию полиморфизма конформации одноцепочечной ПЦР для анализа микробного сообщества на основе гена 16S рРНК. Заявл. Environ. Microbiol. 64, 4870–4876.
Pubmed Реферат | Pubmed Полный текст
Смит, А. Дж. Э. (1978). Моховая флора Великобритании и Ирландии .Кембридж: Издательство Кембриджского университета.
Судзиловская, Н. А., Корнелиссен, Дж. Х. К., Во время, Х. Дж., Ван Логтестейн, Р. С. П., Ланг, С. И., и Аэртс, Р. (2010). Сходная катионообменная способность у видов мохообразных опровергает предполагаемый механизм подкисления торфяников. Экология 91, 2716–2726.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Whinam, J., и Copson, G. (2006). Sphagnum Мох: индикатор изменения климата в субантарктике. Polar Rec. 42, 43–49.
CrossRef Полный текст
Йегер, К. М., Корноски, Л., Хаусман, К., Грот, Э. Э., Белнап, Дж., И Куске, К. Р. (2004). Структура и функционирование диазотрофных сообществ на двух последовательных стадиях биологических почвенных корок плато Колорадо и пустыни Чиуауа. Заявл. Environ. Microbiol. 70, 973–983.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Zachow, C., Тилчер Р. и Берг Г. (2008). Связанные с сахарной свеклой бактериальные и грибковые сообщества демонстрируют высокий местный антагонистический потенциал против патогенов растений. Microb. Ecol. 55, 119–129.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Зер Дж. П. и Тернер П. Дж. (2001). «Фиксация азота: гены нитрогеназ и экспрессия генов», в Methods in Microbiology , ed. Дж. Х. Пол (Нью-Йорк: Academic Press), 271–285.
границ | Редкие виды изменяют структуру бактериальных сообществ в сфагновых компартментах субальпийского торфяника
Введение
Торфяники — это самый крупный наземный резервуар углерода и важный резервуар азота, занимающий площадь 4 × 10 6 км 2 на Земле (Limpens et al., 2008). В глобальном масштабе северные торфяники хранят 473 ~ 621 Гт C и 8 ~ 15 Гт N, что составляет примерно 33% органического углерода почвы и 9 ~ 16% наземного азота (Limpens et al., 2006; Yu et al., 2010) соответственно. Эти обильные органические вещества в основном происходят из мхов Sphagnum , составляющих около 300 видов по всему миру (Daniels, 1990; Shaw et al., 2003). Sphagnum Мхи являются преобладающей растительностью на кислых торфяниках и вносят большой вклад в развитие торфяников. Sphagnum мхи являются хозяевами разнообразных бактериальных сообществ, эндосимбиотических или эктосимбиотических с растением (Opelt et al., 2007a, b; Bragina et al., 2012, 2014; Tian et al., 2019). Бактериальные сообщества могут продуцировать противогрибковые соединения для улучшения защиты хозяев от патогенов и обеспечивать азотом Sphagnum мхов через азотфиксирующие бактерии, такие как Burkholderia (Opelt et al., 2007c; Shcherbakov et al., 2013, 2015). Помимо вышеупомянутых функций, эндосимбиотические метанотрофы, как показано, обеспечивают углекислый газ для фотосинтеза мхов Sphagnum посредством окисления метана (Raghoebarsing et al., 2005; Larmola et al., 2010; Кип и др., 2011). Следовательно, эти микроорганизмы, ассоциированные с Sphagnum , играют важную роль в росте Sphagnum и круговороте элементов в экосистемах торфяников (Bragina et al., 2014; Weston et al., 2015; Kostka et al., 2016).
Как правило, микробные сообщества торфяников состоят из нескольких доминирующих таксонов (относительная численность> 1%) с длинным хвостом видов с низкой численностью (Gilbert, Mitchell, 2006; Andersen et al., 2013; Tian et al., 2019), а также в других местообитаниях (Delgado-Baquerizo et al., 2018). Эти многочисленные таксоны в значительной степени способствуют круговороту углерода и потоку энергии благодаря своей широкой нише, высокой конкурентоспособности и быстрым темпам роста (Campbell et al., 2011). Однако мало известно о популяциях с низкой численностью, обозначенных как редкие биосферные (Sogin et al., 2006), или условно-патогенные виды (Jousset et al., 2017) с высоким разнообразием. Они могут быть частью микробного «семенного банка» и вносить непропорциональный вклад в разнообразие и разнообразие сообществ (Pedrós-Alió, 2007; Shade et al., 2014). Они могут стать доминирующими и выполнять важные функции в круговороте питательных веществ, которые компенсируют функциональный дефицит многочисленных видов (Jousset et al., 2017) в благоприятных условиях окружающей среды (Pedrós-Alió, 2007; Aanderud et al., 2015). Кроме того, реакция многочисленных и редких сообществ на факторы окружающей среды различна (Chen et al., 2017; Ruiz-González et al., 2019) из-за различий в ширине ниши, конкурентоспособности ресурсов и темпах роста (Wu et al., 2017; Xue et al., 2018). Предыдущие исследования микробных сообществ, ассоциированных с Sphagnum , в основном сосредоточены на многочисленных таксонах, тогда как распределение редких видов между различными компартментами Sphagnum (поверхностный торф, эктосфера и эндосфера Sphagnum ) все еще остается в значительной степени неизвестным. Таким образом, мы предполагаем, что (1) редкие виды вносят основной вклад в вариации бактериального сообщества в разных компартментах Sphagnum palustre ; (2) редкие виды могут играть важную роль во всей сети совместного возникновения и потенциальных функциях бактериальных сообществ; (3) редкие виды и многочисленные таксоны могут по-разному реагировать на факторы окружающей среды в поверхностном торфе.
Чтобы проверить наши гипотезы, бактериальные сообщества были исследованы с помощью секвенирования гена 16S рРНК из различных компартментов S. palustre ( торфа S. palustre , эктосферы S. palustre и эндосферы S. palustre ) в Дацзюху. Торфяники, Центральный Китай. В настоящем исследовании мы стремимся ответить на следующие вопросы: (1) сравнить разнообразие и состав многочисленных и редких видов и их относительный вклад в изменение целых бактериальных сообществ; (2) выявить модели совместного присутствия и потенциальные функциональные вариации многочисленных и редких видов в компартментах Sphagnum ; (3) изучить реакцию обильных и редких видов на факторы окружающей среды в S.palustre торф. В целом, это исследование расширит наше понимание бактериальных сообществ, ассоциированных с Sphagnum , в различных компартментах, с упором на редкие виды.
Материалы и методы
Изображение участка и отбор проб
Дацзюху (31 ° 24 ‘- 31 ° 33’ с.ш., 109 ° 56 ‘- 110 ° 11’ в.д.) — типичный субальпийский торфяник, расположенный в Шеннунцзя, провинция Хубэй, центральный Китай (дополнительный рисунок S1A). Торф образовался в закрытой межгорной котловине (1730 м над уровнем моря), площадью около 16 км 2 и глубиной 2–3 м (Huang et al., 2018). Среднегодовая температура и осадки составляют 7,2 ° C и 1560 мм соответственно. Существующие типы растительности — это в основном Carex spp., S. palustre , Sanguisorba officinalis и Euphorbia esula , сопутствующие кустарники. Более подробную информацию о торфяниках Дадзюху предоставили Хуанг и др. (2012) и Ли и др. (2016).
В данном исследовании S. palustre и образцы нижележащего торфа (0–5 см) были собраны с помощью стерильных пластиковых пакетов и центрифужных пробирок соответственно.Образцы были отобраны в трех экземплярах в июле 2016 года. Участки отбора проб были расположены в центральной части торфяника: Эрхаоба (EHB), Нянньянфен (NNF) и два участка в Янлучанге (YLC), где активно росло S. palustre (дополнительный рисунок S1A). ). Уровень грунтовых вод на четырех участках был определен in situ во время отбора проб. Видимые корни растений, опад и остатки были удалены из образцов торфа S. palustre . Все образцы были доставлены в лабораторию геомикробиологии Китайского университета геолого-геофизических исследований (Ухань) в ледяной камере в течение 12 часов.
Обработка образцов
Чтобы удалить микробы из эктосферы, S. palustre промывали трижды по 5 мин каждый раз стерильной деионизированной водой для удаления прилипшего мусора. Затем ок. 5 г образцов S. palustre помещали в 20 мл стерильного фосфатно-солевого буфера (PBS), встряхивали и обрабатывали ультразвуком, чтобы отделить плотно приставшие микробы (Bulgarelli et al., 2015; Edwards et al., 2015). Растительный материал S. palustre переносили в другую пробирку, раствор PBS центрифугировали при 10 000 g в течение 5 минут, а супернатант отбрасывали (дополнительный рисунок S1B).Оставшуюся фракцию PBS хранили при -80 ° C до экстракции ДНК.
Извлеченный S. palustre , предназначенный для эндосферы, подвергся дальнейшей обработке ультразвуком и дважды встряхиванию, следуя описанному выше подходу. Затем образцы S. palustre помещали в 75% этанол и 20% перекись водорода на 1 мин соответственно (Bulgari et al., 2009). Образцы S. palustre промывали три раза по 5 мин каждый раз стерильной деионизированной водой, и конечную стерильную воду инокулировали на среду R2A.Растительный материал S. palustre хранили при -80 ° C до экстракции ДНК.
Дополнительные образцы торфа S. palustre (около 20 г) сушили при 50 ° C в течение 24 часов и подвергали анализу на содержание органического вещества и общего азота.
Экстракция, амплификация и секвенирование ДНК
Суммарная ДНК для каждого приблизительно 0,5 г лиофилизированного S. palustre фракции торфа, эктосферы и эндосферы экстрагировали с помощью набора для выделения ДНК PowerSoil (QIAGEN, Дюссельдорф, Германия).Концентрацию и качество ДНК проверяли с помощью спектрофотометра NanoDrop 2000 (Thermo Fisher Scientific, Уолтем, Массачусетс, США) в 1% -ном агарозном геле, окрашенном бромистым этидием. Области V3-V4 бактериального гена 16S рРНК были амплифицированы с помощью набора праймеров 347F (5′-CCTACGGRRBGCASCAGKVRVGAAT-3 ‘) и 802R (5′-GGACTACNVGGGTWTCTAATCC-3’) (Ren et al., 2019). Реакции ПЦР содержали 25 мкл 2х Premix Taq (Takara Biotechnology, Dalian Co., Ltd., Китай), по 1 мкл каждого праймера (10 мМ) и 3 мкл матрицы ДНК (20 нг / мкл) в объеме 50 мкл.Затем продукты ПЦР очищали и количественно определяли с помощью флуорометра Quit 2.0 (Invitrogen, Карлсбад, Калифорния, США). Библиотеки ДНК были созданы с использованием набора NEBNext Ultra DNA Library Prep Kit для Illumina (New England Biolabs, Ипсвич, Массачусетс, США) в соответствии с протоколами производителя, а качество библиотек было подтверждено с помощью биоанализатора Agilent 2100 (Agilent Technologies, Пало-Альто, Калифорния, США). Штаты) и флуорометр Qubit 2.0. Библиотеки ДНК секвенировали на платформе Illumina MiSeq с считыванием парных концов PE300 в GENEWIZ, Inc.(Сучжоу, Китай).
Анализ последовательности
сырых последовательностей обрабатывали на платформе Quantitative Insight Into Microbial Ecology (QIIME v. 1.9.1) (Caporaso et al., 2010). Последовательности были назначены каждому образцу на основе их уникального штрих-кода с последующим удалением праймеров и штрих-кодов. Прямая и обратная последовательности представляли собой сборку парных концов с перекрытием более 20 п.н. Считывание высокого качества было определено в соответствии с отсутствием неоднозначной базы «N», длиной> 200 п.н. и базовой оценкой качества> 20.Чистые последовательности были сгруппированы в операционные таксономические единицы (OTU) UCLUST (Edgar, 2010) на основании 3% несходства последовательностей, а синглтоны (OTU только с одной последовательностью во всех выборках) были исключены. Митохондрии и хлоропласт были удалены, а химерные последовательности были идентифицированы с помощью алгоритма UCHIME (Edgar et al., 2011). Репрезентативные последовательности для каждой OTU были классифицированы с использованием классификатора Ribosomal Database Project с порогом 0,8 (Cole et al., 2009) и аннотированы в базе данных SILVA 119.Все образцы были повторно проанализированы до 29 577 последовательностей на образец с помощью mothur (Schloss et al., 2009).
Необработанные данные гена 16S рРНК были доступны в базе данных NCBI Sequence Read Archive с регистрационным номером PRJNA512496.
Определение классификации OTU
Мы установили пороги относительной численности на уровне 0,01% для редких таксонов (RT) и 1% для массовых таксонов (AT) и классифицировали все OTU (за исключением архей и неклассифицированных OTU) на шесть подкатегорий (Dai et al., 2016; Xue et al., 2018), чтобы сравнить роли таксонов с разной численностью во всех бактериальных сообществах (WBC): всегда многочисленные таксоны (AAT) с относительной численностью> 1% во всех выборках; условно массовые таксоны (CAT) с относительной численностью ≥1% в некоторых пробах и всегда ≥0.01% во всех пробах; всегда редкие таксоны (ART) с относительной численностью <0,01% во всех пробах; условно редкие таксоны (CRT) с относительной численностью <0,01% в некоторых образцах, но никогда не встречающейся в изобилии (≥1%) в каких-либо образцах; умеренные таксоны (MT) с относительной численностью от 0.01% и 1% во всех выборках; условно многочисленные и редкие таксоны (CART) с относительной численностью от редких (<0,01%) до массовых (≥1%). Подробные описания категории OTU были перечислены в дополнительной таблице S1.
Обработка и анализ даты
Вклад каждой подкатегории в вариации сообщества был определен анализом процента сходства (SIMPER) в PAST 3.26. Индексы альфа-разнообразия (наблюдаемые виды, Chao1, ACE, Simpson, Pielou и Shannon), NMDS, PERMANOVA и ANOSIM были выполнены с использованием «веганского» пакета в R 3.5.1. Функциональные профили были спрогнозированы с использованием пакета Tax4Fun (Aßhauer et al., 2015), а анализ главных компонентов (PCA) был использован для выявления компартментных различий потенциальных функциональных сообществ на основе евклидова расстояния относительной численности общей Киотской энциклопедии генов и геномов (KEGG). ) Ортологи (КО) с пакетом «ggord» в R 3.5.1. Различия в альфа-разнообразии и потенциальной функции были проверены с помощью одностороннего дисперсионного анализа (ANOVA) в SPSS 18. Анализ бета-разнообразия был выполнен на основе расстояния Брея-Кертиса относительной численности OTU в неметрическом многомерном масштабировании (NMDS).Анализ сходства (ANOSIM) использовался для проверки различий между группами. Тесты Монте-Карло анализа избыточности определили взаимосвязь между факторами окружающей среды и бактериальными сообществами торфа S. palustre в Каноко 5.
Были отобраны OTU с относительной численностью выше 0,01%, и корреляционная матрица между двумя OTU была рассчитана в пакете «mental» R. Коэффициент корреляции ( r > 0,8 или r <-0,8) и значимость (с поправкой на FDR P <0.01) были интегрированы в сетевой анализ (Jiao et al., 2016). Атрибуты топологии сети (количество узлов и ребер, средняя степень, диаметр сети, плотность графа, модульность, коэффициент кластеризации и средняя длина пути) были рассчитаны с использованием пакета «igraph» в R 3.5.1. Визуализация сети проводилась с помощью Gephi 0.9.2. Тем временем было построено 1000 случайных сетей Эрдеша-Рейни для сравнения с реальной сетью (Erdös and Réyni, 1960). Узлы с высокой степенью (> 100) и низкой промежуточной центральностью (<5000) считались ключевыми видами в сетях совместного возникновения (Ma et al., 2016).
Результаты
Разнообразие бактериальных сообществ
Всего было получено 2 413 649 последовательностей высокого качества и 3144 операционных таксономических единиц (OTU). После удаления архей и неклассифицированных OTU 2 848 OTU были подвергнуты дальнейшему анализу. Среди них 13 OTU, представляющих 9,01% всех последовательностей, были многочисленными таксонами (AT) и присутствовали во всех выборках, тогда как 2706 OTU, занимающих 39,2% всех последовательностей, принадлежали к редким таксонам (RT). Условно обильные и редкие таксоны (CART) включали 126 OTU, представляющих 51.13% всех последовательностей (дополнительная таблица S1).
Значительные различия наблюдались в альфа-разнообразии целых бактериальных сообществ (WBC) и RT между компартментами (дополнительная таблица S2). Наблюдаемые виды, индексы Симпсона, Пиелу и Шеннона WBC и CART значительно снизились с Sphagnum palustre торфа (SP) до эндосферы S. palustre (Endo), с таковыми для эктосферы S. palustre (Ecto) в г. между (Рисунок 1). Напротив, не было значительных различий в альфа-разнообразии на разных участках отбора проб (дополнительная таблица S2).Диаграммы Венна показали, что более 50% OTU в WBC были разделены в трех отсеках, а распределение общих и специфических OTU в RT было аналогично таковому для WBC (Рисунок 2B).
Рис. 1. Изменения в альфа-разнообразии (наблюдаемые виды, индексы Симпсона, Пиелу и Шеннона) в компартментах Sphagnum palustre . Односторонний дисперсионный анализ (ANOVA) используется для сравнения различий между компартментами S. palustre . Крестик представляет собой среднее значение ( n = 12).WBC, целые бактериальные сообщества; AT — многочисленные таксоны; RT — редкие таксоны; CART, условно обильные и редкие таксоны; SP, S. palustre торф; Ecto, S. palustre эктосфера; Endo, S. palustre эндосфера. Значения F и P наблюдаемых видов не могут быть рассчитаны в AT. Цветные точки представляют образцы SP, Ecto и Endo соответственно. Линии — это подогнанные кривые.
Рис. 2. Структурные вариации бактериальных сообществ S.palustre отсеков. (A) Структура бактериальных сообществ на основе расстояния Брея-Кертиса в NMDS. (B) Количество общих и уникальных OTU в разных компартментах S. palustre . (C) Различия сообществ бактериальных сообществ между компартментами S. palustre . Каждый прямоугольник указывает максимальный, минимальный, 75-й и 25-й квартили соответственно, а линия в каждом прямоугольнике представляет собой медианное значение ( n = 144).Строчные буквы указывают на значения при 95% доверительном интервале согласно ANOVA с апостериорным тестом HSD Тьюки. Аббревиатуры такие же, как показано на Рисунке 1.
Различия в бактериальных сообществах по компартментам визуализировали с помощью анализа NMDS на основе матрицы Брея-Кертиса. Сообщества одного компартмента сгруппированы вместе (рис. 2А). Попарное сравнение межгрупповых расстояний (рис. 2C) и результаты ANOSIM (таблица 1) показали, что бактериальные сообщества значительно различались между компартментами, а не между участками отбора проб.
Таблица 1. Тест на несходство, показывающий различия бактериальных сообществ по компартментам и участкам отбора проб.
Состав бактериальных сообществ
Что касается лейкоцитов, то в SP преобладали ацидобактерии и альфапротеобактерии, тогда как преобладающими типами в Ecto были цианобактерии, альфапротеобактерии и гаммапротеобактерии, за которыми следовали ацидобактерии. В Endo в сообществах преобладали цианобактерии и Alphaproteobacteria, за которыми следовали ацидобактерии (рис. 3A).Alphaproteobacteria обычно обнаруживались во всех компартментах, а Acidobacteria часто присутствовали в SP, тогда как Cyanobacteria в изобилии присутствовали в Ecto и Endo.
Рисунок 3. Состав бактериальных сообществ в WBC (A) , AT (B) , RT (C) и CART (D) . Е1, первое место Эрхаобы; N1, первое место Niangniangfen; Y2, второй участок Янлучана; Y6, шестой участок Янлучан. Другие сокращения показаны на Рисунке 1.
Всего 13 OTU были классифицированы в AT, которые были связаны с Actinobacteria, Alphaproteobacteria и Gammaproteobacteria phyla (рис. 3B), и на их долю приходилось 0,46% от богатства OTU (дополнительная таблица S3). Целых 2706 OTU были идентифицированы как RT, которые были связаны с 30 типами (рис. 3C) и представляли 95,01% богатства OTU (дополнительная таблица S3). Кроме того, 126 OTU были признаны CART, принадлежащими 13 типам (рис. 3D), и представляли 4,42% от богатства OTU (дополнительная таблица S3).
Сеть совместного возникновения
Сеть совместной встречаемости была построена на основе корреляционных отношений между относительной численностью OTU. Полученная сеть содержала 674 узла, соединенных 8 455 ребрами, где обычно наблюдались положительные корреляции по сравнению с отрицательными (рис. 4C). Безмасштабное распределение степеней реальной сети (степенной закон: R 2 = 0,945, P <0,0001, дополнительный рисунок S2) указывает на безмасштабную и неслучайную структуру сети.Между тем сеть имела особенности малого мира и модульную структуру, на что указывают более высокие значения индекса модульности (0,762) и коэффициента кластеризации (0,521) по сравнению со случайными сетями Эрдеша-Рейни одинакового размера (Erdös and Réyni, 1960; Таблица 2). .
Рис. 4. Сеть совместного присутствия бактериальных сообществ, отсортированная по цвету по типу (A) , модульности (B) и категории OTU (C) ранга соответственно. Связь между двумя узлами означает значительную корреляцию ( r > 0.8 или r <–0,8, P <0,01). Цифры в круглых скобках указывают количество узлов в (A, B, C) . Каждый узел представляет собой одну OTU. Положительные и отрицательные номера ребер среди таксонов с разной численностью помещены снаружи и внутри скобок на небольшой панели в правом нижнем углу. Размер каждого узла пропорционален его степени; толщина кромки, соединяющей два узла, пропорциональна соответствующему коэффициенту корреляции. RT — редкие таксоны; CART, условно обильные и редкие таксоны; AT — многочисленные таксоны; MT, умеренные таксоны.
Таблица 2. Топологические свойства сетей совместного возникновения микробных сообществ на торфяниках Дайзюху.
На 10 основных типов сети приходилось 93% всех узлов (рис. 4A). Вся сеть была сгруппирована в 4 основных модуля (рис. 4B). Модуль I включал 36,35% узлов всей сети, которые в основном попали в Proteobacteria. Модуль II и модуль III в основном состояли из Proteobacteria и Acidobacteria, занимающих 41 место.99% узлов во всей сети, тогда как модуль IV в основном состоял из Proteobacteria и Bacteroidetes (дополнительный рисунок S3A). Трехкомпонентные графики показали специфичность различных модулей к определенному компартменту (дополнительные рисунки S3B – E). Например, большинство OTU в модуле I показали высокие доли в Ecto и Endo, тогда как большинство OTU в модулях II, III и IV продемонстрировали высокие пропорции в SP.
Топологические особенности на уровне узлов микробных групп с разной численностью (RT, CART, AT и MT) показали, что значения степени, промежуточности и центральности собственного вектора для CART были значительно выше, чем у RT (дополнительные рисунки S4A, B, D) .Значения центральности близости AT были самыми высокими среди 4 микробных групп (дополнительный рисунок S4C). На основании высокой степени (> 100) и низкой промежуточности (<5000) в сетях совместного возникновения 5 OTU были идентифицированы как ключевые виды, включая Acidobacteria (1 OTU), Betaproteobacteria (1 OTU), Alphaproteobacteria (1 OTU), Ignavibacteriae (1 OTU) и Spirochaetae (1 OTU), которые принадлежали RT и CART (дополнительная таблица S4).
Факторы окружающей среды, связанные с бактериальными сообществами
Тесты Монте-Карло показали, что три физико-химических параметра, т.е.е. уровень грунтовых вод, содержание общего азота и органических веществ в значительной степени коррелировали с вариациями WBC, RT, CART и AT в образцах SP (Таблица 3). Уровень лейкоцитов значительно зависел от уровня грунтовых вод и общего содержания азота ( P <0,05), которые также были определены как значимые детерминанты ( P <0,05) в RT и CART. Напротив, содержание органических веществ, а не уровень грунтовых вод и общий азот значительно ( P <0,05) сформировали структуру AT.
Таблица 3. Корреляция между бактериальными сообществами и факторами окружающей среды в образцах торфа S. palustre на торфяниках Дайзюху.
Возможные функции бактериальных сообществ
Подобно структурным вариациям сообществ, потенциальные функции сообществ продемонстрировали очевидное разделение между WBC, RT и CART (дополнительные рисунки S5A, C, D) по сравнению с таковыми в AT по компартментам (дополнительный рисунок S5B), как указано основными компонентный анализ (PCA) на основе евклидова расстояния.Сравнения различий в каждых двух компартментах и участках отбора проб были дополнительно количественно оценены с использованием PERMANOVA (дополнительная таблица S5), которая показала значительные (α = 0,05) различия функциональных потенциалов между отделами, а не между участками отбора проб.
Среди WBC, RT и CART относительное количество потенциальных азотфиксирующих функциональных групп было самым высоким у SP, за ним следовали группы Endo, а самым низким было у Ecto (рис. 5A). Относительное содержание предсказанных функциональных групп, окисляющих метан, значительно снизилось от Endo к SP, с промежуточным содержанием Ecto (Рисунок 5B).Вместе определенные потенциальные функциональные группы, такие как N 2 -фиксаторы и CH 4 -окислители в RT и CART, могут отражать согласованные паттерны вариаций с таковыми в WBC по компартментам, тогда как AT нет.
Рис. 5. Относительное содержание предсказанных азотфиксирующих (A) и метанокисляющих групп (B) в WBC, AT, RT и CART. Сокращения те же, что и на рисунке 1. Значимые различия (тесты ANOVA) обозначены следующим образом: ∗∗ P <0.01; ∗∗∗ P <0,001. Строчные буквы обозначают значимые различия при доверительном интервале 95%.
Обсуждение
Основная цель этого исследования заключалась в изучении роли таксонов с различной численностью в сдвиге бактериальных сообществ в компартментах Sphagnum на субальпийском торфянике. Наши результаты показали, что редкие виды (относящиеся к RT и CART и далее) были основными участниками перемещения бактериальных сообществ, ассоциированных с Sphagnum , в различные компартменты.Редкие виды продемонстрировали постоянную изменчивую структуру с WBC в структурах, предсказали функциональные профили и реакцию на факторы окружающей среды. Кроме того, они оказали большое влияние на сеть совместной встречаемости WBC. Таким образом, наше исследование позволяет по-новому взглянуть на роль редких видов в микробных сообществах, связанных с Sphagnum .
Изменения в бактериальных сообществах
Наши результаты подтвердили, что эффекты компартментов привели к сдвигу бактериальных сообществ в S.palustre как указано альфа- и бета-разнообразием. Альфа-разнообразие снизилось от SP до Ecto и было самым низким в Endo (Рисунок 1). Этот результат согласуется с точкой зрения на микробную колонизацию, связанную с растениями, которая показывает сокращение разнообразия видов от ризосферы до ризопланы и эндосферы (Edwards et al., 2015; Beckers et al., 2017). Изменения альфа-разнообразия от SP к Ecto к Endo могут быть тесно связаны со специфическими тканями Sphagnum , состоящими из хлороцитов и гиалоцитов.Микробная колонизация в Sphagnum строго отбирается хлороцитами (Opelt et al., 2007b; Bragina et al., 2012) и снижает бактериальное разнообразие в компартментах Sphagnum (Рисунок 1). Значительное увеличение различий в сообществах (бета-разнообразие) между компартментами от Ecto против Endo до SP против Ecto и далее до SP против Endo (Рисунок 2C и Таблица 1) также подтвердило влияние компартмента растений на бактериальные сообщества. Каждый отсек представляет собой уникальную и гетерогенную микросреду (Coleman-Derr et al., 2016; Stacie et al., 2016), который выбирает их конкретные микробные сообщества (Beckers et al., 2017). Эктосфера растений обычно олиготрофна с частыми колебаниями температуры, осадков, влажности и радиации, что затрудняет выживание микробов (Müller et al., 2016). Поэтому микроорганизмам, живущим на поверхности растений, приходится иметь дело с жесткими условиями, что привело к дисперсному распределению бактериальных сообществ в Ecto (рис. 2A). Напротив, клетки растений могут действовать как оазисы для выживания микробов на торфяниках.Обычно микроорганизмы со специфическими признаками, например хемотаксисом, подвижностью и восприятием кворума, привлекаются в результате зависимого от хозяина отбора или пассивного транспорта и, таким образом, приводят к образованию набора специфических связанных с растением микробных сообществ (Compant et al., 2010; Kardol, Wardle, 2010; Bulgarelli et al., 2012; Philippot et al., 2013; Müller et al., 2016). В отличие от сосудистых растений, безкорневые мхи Sphagnum перехватывают и удерживают воду и питательные вещества через нефотосинтезирующие клетки и соседние фотосинтетические клетки (Weston et al., 2015; Kostka et al., 2016), что может облегчить захват их эндосимбиотических микроорганизмов.
Наши результаты впервые показали, что таксоны микробов с разной численностью объясняют различные вариации микробных сообществ на субальпийском торфянике. Богатство RT было более чем в 60 раз выше, чем у AT (Рисунок 1), что свидетельствует об их важном вкладе в микробное разнообразие (Lynch and Neufeld, 2015; Zhan and Macisaac, 2015). Паттерны изменчивости микробных таксонов с разной численностью альфа- и бета-разнообразия среди S.palustre (рис. 1, 2) подтверждают гипотезу 1 о том, что редкие виды в основном вносят вклад в вариации бактериального сообщества по сравнению с многочисленными таксонами.
Схема совместного возникновения сообществ в Сфагнум Отделения
Результаты сетевого анализа совместной встречаемости показали, что ключевые виды имеют решающее значение для всего бактериального сообщества и могут оказывать большое влияние на другие компоненты сообщества. Сеть совместного возникновения может обеспечить глубокое и уникальное понимание микробных взаимодействий и собраний сообщества (Barberán et al., 2012). Наши результаты показали, что между RT и CART наблюдалось больше положительных взаимодействий (рис. 4C), чем отрицательных, что предполагает возможное сотрудничество между ними (Ju et al., 2014). Краеугольные виды характеризуются высокой степенью и низкой промежуточностью центральности, основанной на безмасштабном признаке (Barabási, 2009; Berry and Widder, 2014), и признаны начальными компонентами и влияют на другие компоненты сообщества в сетях. В нашем исследовании все идентифицированные ключевые виды были связаны с RT и CART, что подтвердило, что редкие виды могут играть решающую роль в сети совместного появления сообществ.Среди ключевых видов Candidatus Nitrotoga — адаптированные к холоду нитритокисляющие бактерии, участвующие в круговороте азота (Alawi et al., 2007). Spirochaeta — анаэробные бактерии, некоторые представители которых способны к разложению производными целлюлозы (глюкоза и целлобиоза) и выработке метана в торфяных почвах (Schmidt et al., 2015; Juottonen et al., 2017). Эти ключевые виды могут способствовать развитию Sphagnum в экстремальных условиях и опосредовать переработку макроэлементов (C, N) (Bragina et al., 2014) и поддерживать экосистемную сеть (Layeghifard et al., 2017). Следовательно, редкие виды могут играть важную роль в поддержании функционирования экосистемы торфяников.
Следует отметить, что редкие виды широко распространены во всех модулях, что может повлиять на функционирование окружающей среды, связанной с хозяином (Jousset et al., 2017). Например, азотфиксирующие и окисляющие метан микробы играют важную роль в поставке азота для Sphagnum (Larmola et al., 2014; Vile et al., 2014) и разложении органических веществ (Chen and Murrell, 2010).В прогнозируемых функциональных сообществах вариации в относительном количестве генов, кодирующих нитрогеназу и метанмонооксигеназу в RT и CART, соответствовали таковым в WBC между компартментами (рис. 5). Таким образом, мы предположили, что редкие виды играют важную роль в сети совместного возникновения и определенных функциях бактериальных сообществ, и гипотеза 2 была поддержана. Хотя сетевой анализ может дать нам некоторое представление о микробных взаимодействиях, он показывает только статистическую корреляцию между микробными группами и не доказывает напрямую микробные взаимодействия.Комбинация экспериментов на основе микроскопии или совместного культивирования с анализом совместной встречаемости предоставит более прямые доказательства микробных взаимодействий.
Факторы окружающей среды, формирующие бактериальные сообщества
Условия окружающей среды сильно влияют на структуру микробных сообществ (Bahram et al., 2018). Влияние уровня грунтовых вод и общего содержания азота на бактериальные сообщества в образцах SP (таблица 3) хорошо согласуется с предыдущими исследованиями на различных типах торфяников (Mishra et al., 2014; Урбанова и Барта, 2016; Чжун и др., 2017). Интересно, что мы также обнаружили, что RT и CART также реагировали на уровень грунтовых вод и общее содержание азота. Напротив, AT был чувствителен к содержанию органических веществ. Это указывает на то, что таксоны микробов с разной численностью по-разному реагируют на факторы окружающей среды. Было продемонстрировано, что содержание и форма органического вещества влияют на биодоступность и качество органического углерода, а также влияют на приоритет микробного метаболизма (Stres et al., 2008). Наши результаты также показали, что содержание органических веществ в основном влияет на АТ в лейкоцитах. Таким образом, была подтверждена гипотеза 3 о том, что редкие виды и многочисленные таксоны по-разному реагируют на факторы окружающей среды.
Группы микробов с разной численностью показали разные реакции окружающей среды, которые могут быть результатом фильтрации или ограничения распространения в окружающей среде (Chen et al., 2017; Wu et al., 2017; Xue et al., 2018). Наши результаты (таблица 3) показали, что RT и CART были более чувствительны к фильтрации окружающей среды, чем AT.Обычно RT и CART демонстрировали слабую конкурентоспособность и низкую скорость роста (Campbell et al., 2011; Logares et al., 2015), тогда как AT может конкурировать за большее количество субстратов и хорошо адаптироваться к экосистеме за счет активного роста и распространения в широкой нише. (Pedrós-Alió, 2006; Jousset et al., 2017; Xue et al., 2018). Это говорит о том, что на состав микробных сообществ сильно влияет сортировка видов, вызванная окружающей средой (Roberto et al., 2018). Таким образом, таксоны с разной численностью в бактериальных сообществах из SP подвергались влиянию неравномерной фильтрации окружающей среды.
ЗАКЛЮЧЕНИЕ
Наши результаты показали, что структуры и прогнозируемые функции редких видов в значительной степени смещены в разных компартментах S. palustre в соответствии с таковыми для целых бактериальных сообществ. Эти результаты расширили наши знания о воздействии компартмента на бактериальные сообщества торфяников, подчеркнув важность редких видов. Сетевой анализ совместной встречаемости показал, что редкие виды могут играть центральную роль в бактериальных сообществах.Как и целые бактериальные сообщества, редкие виды также в значительной степени определялись уровнем грунтовых вод и общим содержанием азота в SP, тогда как многочисленные таксоны более тесно коррелировали с содержанием органического вещества. В совокупности наши результаты подчеркивают важное влияние редких видов на бактериальные сообщества. Редкие виды в основном вносят вклад в различия как в структуре, так и в потенциальных функциях микробных сообществ, связанных с Sphagnum , между компартментами субальпийских торфяных экосистем.
Заявление о доступности данныхНаборы данных, созданные для этого исследования, можно найти в базе данных NCBI Sequence Read Archive (PRJNA512496).
Авторские взносы
WT провел эксперименты, проанализировал данные и подготовил рукопись. ХХ помог анализ данных. LM, SE и XQ дали свои предложения по рукописи. RW помог собрать образцы. HW разработал исследование, предоставил финансирование и отредактировал рукопись.
Финансирование
Это исследование было поддержано Национальным фондом естественных наук Китая (41572325), Фондами фундаментальных исследований для центральных университетов, Китайским университетом геонаук (Ухань) (CUGCJ1703, CUGQY1922) и Фондом естественных наук провинции Хубэй (2019CFB245). ).
Конфликт интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим доктора Байин Мана и доктора Юань Юня за их помощь в процессе эксперимента. Мы также благодарим редактора HC-Q и двух рецензентов за их проницательные комментарии, улучшившие ясность этой статьи.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https: // www.frontiersin.org/articles/10.3389/fmicb.2019.03138/full#supplementary-material
Сноски
Список литературы
Аандеруд, З. Т., Джонс, С. Э., Фирер, Н., Леннон, Дж. Т. (2015). Реанимация редкой биосферы способствует импульсам активности экосистемы. Фронт. Microbiol. 6:24. DOI: 10.3389 / fmicb.2015.00024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Алави, М., Липски, А., Сандерс, Т., Пфайффер, Э. М., и Спик, Э. (2007). Выращивание новой адаптированной к холоду нитритокисляющей бета-протеобактерии из сибирской Арктики. ISME J. 1, 256–264. DOI: 10.1038 / ismej.2007.34
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Андерсен, Р., Чепмен, С. Дж., И Арц, Р. Р. Э. (2013). Микробные сообщества естественных и нарушенных торфяников: обзор. Soil Biol. Biochem. 57, 979–994. DOI: 10.1016 / j.soilbio.2012.10.003
CrossRef Полный текст | Google Scholar
Aßhauer, K. P., Wemheuer, B., Daniel, R., and Meinicke, P. (2015). Tax4Fun: прогнозирование функциональных профилей на основе данных метагеномной 16S рРНК. Биоинформатика 31, 2882–2884. DOI: 10.1093 / биоинформатика / btv287
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бахрам, М., Хильдебранд, Ф., Форслунд, С. К., Андерсон, Дж. Л., Судзиловская, Н. А., Бодегом, П. М. и др. (2018). Структура и функции глобального микробиома верхнего слоя почвы. Природа 560, 233–237. DOI: 10.1038 / s41586-018-0386-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барберан А., Бейтс С. Т., Касамайор Э. О. и Фирер Н. (2012). Использование сетевого анализа для изучения моделей совместной встречаемости в микробных сообществах почвы. ISME J. 6, 343–351. DOI: 10.1038 / ismej.2013.236
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бекерс, Б., Бек, М. О., Вайенс, Н., Боэрджан, В., и Вангронсвельд, Дж. (2017). Структурная изменчивость и дифференциация ниш в ризосферном и эндосферном бактериальном микробиоме полевых деревьев тополя. Микробиом 5, 25–41. DOI: 10.1186 / s40168-017-0241-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берри Д. и Виддер С. (2014). Расшифровка микробных взаимодействий и обнаружение ключевых видов с помощью сетей совместного возникновения. Фронт. Microbiol. 5: 219. DOI: 10.3389 / fmicb.2014.00219
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брагина А., Берг К., Кардинале М., Щербаков А., Чеботарь В., Берг Г. (2012). Sphagnum мхи обладают высокоспецифическим бактериальным разнообразием на протяжении всего своего жизненного цикла. ISME J. 6, 802–813. DOI: 10.1038 / ismej.2011.151
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брагина, А., Обераунерваппис, Л., Захов, К., Халвакс, Б., Таллингер, Г.Г., Мюллер, Х. и др. (2014). Микробиом Sphagnum поддерживает функционирование болотной экосистемы в экстремальных условиях. Мол. Ecol. 23, 4498–4510. doi: 10.1111 / mec.12885
Frontiers | Инактивация и побег X: эпигенетические и структурные особенности
Введение
Эволюция половых хромосом млекопитающих из пары аутосом привела к появлению отдельных гетероморфных хромосом, которые управляют определением пола (Graves, 2016). Y-хромосома содержит несколько генов (∼70) и присутствует только у мужчин, в то время как X-хромосома содержит много генов (∼900–1500) и присутствует в двух копиях у женщин и одной копии у мужчин.Это способствует дисбалансу дозировки генов между Х-сцепленными и аутосомными генами и между полами (Disteche, 2016). Чтобы облегчить этот дисбаланс, были разработаны два механизма дозовой компенсации: активация X экспрессируемых генов у мужчин и женщин и инактивация или подавление X хромосомы у женщин (Deng et al., 2014).
Здесь мы сосредотачиваемся на инактивации Х-хромосомы (XCI), механизме, который приводит к подавлению случайно выбранной Х-хромосомы в раннем женском эмбриогенезе (Lyon, 1961).XCI характеризуется каскадом молекулярных событий, начинающихся вскоре после имплантации эмбриона, и точно поддерживается во всех соматических клетках организма, обеспечивая надежную модель для изучения эпигенетических и структурных изменений, связанных с молчанием генов (Galupa and Heard, 2018). Этот сложный процесс начинается с покрытия cis- будущей неактивной X-хромосомы (Xi) длинной некодирующей РНК (lncRNA) Xist (Borsani et al., 1991; Brockdorff et al., 1991; Brown и другие., 1991). Слои модификаций хроматина и ДНК, катализируемые белками, первоначально рекрутируемыми РНК Xist , затем помещаются на место в течение нескольких дней во время раннего развития для стабильного подавления транскрипции каждого гена на Xi (Froberg et al., 2013; Mira-Bontenbal and Gribnau , 2016). Эти модификации связаны с глубокими изменениями в трехмерной структуре и расположении Xi, оба процесса зависят от локусов X-сцепленной lncRNA. Xi принимает двудольную структуру, состоящую из двух супердоменов конденсации хроматина, разделенных локусом днРНК Dxz4 , и Xi посещает ядрышко, процессу, которому способствует днРНК Firre (Zhang et al., 2007; Рао и др., 2014; Deng et al., 2015; Минаджиги и др., 2015; Ян и др., 2015; Джорджетти и др., 2016; Fang et al., 2019).
Несмотря на множественные уровни репрессии генов, которые стабилизируют XCI, субнабор критических для развития генов остается экспрессируемым, хотя и на более низком уровне, от Xi (Carrel and Willard, 2005; Berletch et al., 2011). Эти ускользающие гены принимают сигнатуры хроматина и структурные особенности, более похожие на те, что обнаруживаются в областях активной транскрипции (Balaton and Brown, 2016).Такие гены могут иметь более высокую экспрессию у женщин, что приводит к половым различиям в нормальной физиологии и в восприимчивости к заболеваниям. Аномальная дозировка гена ускользания способствует возникновению целого ряда вредоносных фенотипов, включая бесплодие, умственную отсталость, иммунные заболевания и рак (Disteche, 2016; Balaton et al., 2018).
Этот обзор фокусируется в первую очередь на механизмах, которые регулируют структуру Х хромосомы и расположение ядра по отношению к XCI, с особым акцентом на роли X-связанных lncRNAs в этих процессах.Затем мы обсудим механизмы, которые позволяют избранному подмножеству генов избегать молчания в контексте репрессированной среды Xi, и то, как секвенирование РНК одной клетки используется для идентификации новых генов ускользания. Наконец, мы рассматриваем новые данные о роли дозы гена ускользания в половых различиях в состоянии здоровья и болезни.
Длинные некодирующие РНК контролируют эпигенетические и структурные особенности неактивной Х-хромосомы
Значение днРНК в контроле ядерной структуры и экспрессии генов становится все более очевидным (Engreitz et al., 2016). Здесь мы рассматриваем три X-сцепленных lncRNAs, Xist , Dxz4 и Firre , которые участвуют в различных аспектах возникновения и поддержания XCI.
Xist
Недавние обзоры подробно рассмотрели роль Xist (Mira-Bontenbal and Gribnau, 2016; Galupa and Heard, 2018). Таким образом, мы сосредоточим нашу дискуссию на вопросах, связанных со структурными изменениями на Си. Одним из них является локальное изменение конформации хроматина в центре инактивации X (XIC), которое важно для правильного инициирования XCI.XIC содержит как локус Xist , так и его антисмысловую транскрипционную единицу Tsix , вместе с множеством других локусов, которые регулируют Xist (Froberg et al., 2013; Galupa and Heard, 2018). Промоторы Xist и Tsix лежат в отдельных, но смежных регионах локальных взаимодействий хроматина, называемых топологически связанными доменами (TADs) (Nora et al., 2012). Интересно отметить, что замена транскрипционной единицы Xist / Tsix и размещение их промоторов в TAD друг друга приводит к переключению в динамике их экспрессии, указывая на то, что топологическое разделение XIC имеет решающее значение для правильной инициации XCI (van Bemmel et al., 2019).
Как только инициируется XCI, структурные изменения на всей хромосоме приводят к образованию конденсированных тельцов Барра, покрытых Xist RNA (Figure 1). Визуализирующие исследования показывают быстрое формирование ядерного компартмента, лишенного транскрипционного аппарата и эухроматических меток, в котором Х-сцепленные гены, изначально расположенные на периферии облака РНК Xist , занимают более внутреннее положение при замалчивании (Chaumeil et al. , 2006; Клемсон и др., 2006). Последующий анализ конформации хроматина с помощью Hi-C демонстрирует, что Xist важен для образования уникальной двудольной структуры Xi, которая дополнительно обсуждается ниже (см. Раздел « Dxz4 ») (Minajigi et al., 2015; Джорджетти и др., 2016). Большая часть Xi показывает ослабление локальных TAD и больших A / B-компартментов активного и неактивного хроматина, что обычно проявляется на аутосомах и активной X-хромосоме (Lieberman-Aiden et al., 2009; Dixon et al., 2012; Minajigi et al. др., 2015; Джорджетти и др., 2016). Xist RNA pull-down исследования идентифицировали два РНК-связывающих белка, участвующих в фазовом переходе жидкость-твердое тело, FUS и hnRNPA2, предполагая возможность того, что разделение фаз способствует образованию гетерохроматина и сайленсингу Xi (Calabrese et al., 2012; Patel et al., 2015; Райан и др., 2018). Действительно, предполагается, что большинство белков в интерактоме Xist склонны к фазовому разделению. С помощью РНК-FISH высокого разрешения можно идентифицировать около сотни фокусов Xist на Xi, причем эти фокусы имеют сравнимую форму, размер и морфологию с другими фазово-разделенными конденсатами, такими как параспеклы и стрессовые гранулы (Cerase et al., 2019).
Рисунок 1. Длинные некодирующие РНК контролируют эпигенетические и структурные характеристики Xi. (A) XCI начинается с выражения Xist , расположенного в XIC (черный). Xist РНК распространяется (волнистая черная линия) вдоль Х-хромосомы и рекрутирует несколько белковых комплексов (см. Текст). Происходят основные модификации гистонов, включая деацетилирование гистонов с помощью HDAC3, за которым следует убиквитинирование h3AK119, опосредованное комплексом PRC1, метилирование h4K27, опосредованное комплексом PRC2, и, наконец, включение гистона macroh3A. Ген A представляет собой ген, который становится инактивированным (красный), в то время как ген B является геном ускользания (зеленый), который остается неизменным.Эта схема фокусируется на модификациях гистонов и не показывает рекрутирование других белков или других репрессивных эпигенетических модификаций, таких как метилирование ДНК. Под схемой показаны примеры ядер самок мышей, показывающих облако Xist (зеленый) после РНК-FISH и обогащение модификаций гистонов (розовый) путем иммуноокрашивания h3AK119Ubi, h4K27me3 и macroh3A. Ядра контрастируют с помощью Hoechst 33342. (B) Геномное расположение Firre , Dxz4 и Xist указано на схеме X-хромосомы мыши вдоль контактной карты Hi-C Xi мыши Patski клетки.Два супердомена частых контактов разделены областью Dxz4 . Цветовая шкала показывает нормализованное количество контактов [адаптировано из опубликованного рисунка (Bonora et al., 2018) в Nature Communications под лицензией Springer Nature Publishing License: http://creativecommons.org/licenses/by/4.0/. (C) Предпочтительные местоположения Xi находятся рядом с ядрышком или на периферии ядра, как показано на примерах ядер фибробластов мыши после РНК-FISH для Xist (зеленый) для определения местоположения Xi и иммуноокрашивания на нуклеофозмин (красный) для определения местоположения ядрышко.Ядра контрастируют с помощью Hoechst 33342.
Xist РНК-опосредованное молчание генов достигается за счет рекрутирования белков, которые устанавливают эпигенетические и структурные модификации на Xi (рис. 1A; Chu et al., 2015; McHugh et al., 2015; Minajigi et al., 2015; Moindrot) et al., 2015; Monfort et al., 2015). Первоначально распространение РНК Xist и ее белковых комплексов считалось линейным, но новые исследования выявили сложные взаимодействия между эпигенетическими и геномными особенностями, такими как геномное расстояние от локуса Xist , плотность генов и близость к давно перемежающимся объектам. ядерные элементы (LINE), которые действуют как путевые станции для усиления покрытия РНК Xist по всему пространству Xi 3D (Sousa et al., 2019). Анализ последовательности РНК Xist показывает, что значительная часть последовательности первичной РНК состоит из блоков локальных тандемных повторов (названных A-F) с различными функциями в XCI. Первостепенное значение A-повтора в подавлении молчания X, но не в покрытии РНК Xist было признано на раннем этапе и было дополнительно подтверждено демонстрацией того, что он рекрутирует репрессор транскрипции SPEN (Wutz et al., 2002; Zhao et al. , 2008; Нестерова и др., 2019). B-повтор вместе с короткой частью C-повтора имеют решающее значение для распространения РНК Xist и для привлечения поликомбинированных комплексов сайленсинга PRC1 и PRC2 (Pintacuda et al., 2017; Нестерова и др., 2019). Напротив, устранение PRC1 или PRC2 нарушает распространение Xist , поддерживая интерактивные роли этого комплекса мега-белок-РНК (Colognori et al., 2019).
Новое исследование теперь прояснило точный порядок появления модификаций гистонов по отношению к замалчиванию X (Żylicz et al., 2019). Важно отметить, что потеря ацетилирования гистонов, в частности h4K27ac, несомненно, является одним из самых первых событий после накопления РНК Xist во время инициации XCI (Рис. 1A).В самом деле, гистондеацетилаза HDAC3 предварительно загружена в предполагаемые энхансеры и жизненно важна для эффективного подавления большинства генов на Xi. Убиквитинирование гистона h3K119 затем инициируется комплексом ядерный матрикс-белок PRC1 (hnRNPK-PCGF3 / 5-PRC1), сигнализируя о последующем рекрутировании других комплексов PRC1 и PRC2 (рисунки 1A, B; Pintacuda et al., 2017). Триметилирование гистона h4K27 опосредуется PRC2 и появляется несколько позже, даже после подавления гена (Żylicz et al., 2019). Каркасный белок хроматина SMCHD1 (структурное поддержание хромосомного гибкого шарнирного домена, содержащего белок 1) играет важную роль в подавлении генов и структуре Xi (Blewitt et al., 2008). Недавнее исследование предполагает, что двудольная структура Xi формируется через промежуточную стадию конденсации, опосредованную SMCHD1, в которой A / B-компартменты изначально сливаются в S1 / S2-компартменты, которые впоследствии сливаются в безкомпартментную архитектуру Xi (Wang et al., 2018 ). В соответствии с этим открытием, потеря функции SMCHD1 приводит к появлению субмегабазных доменов, A / B-компартментов и частичному восстановлению границ TAD на Xi (Gdula et al., 2019). Эти изменения связаны с дерепрессией некоторых X-инактивированных генов и локальным уменьшением h4K27me3, но эта реактивация не наблюдается в иммортализованных эмбриональных фибробластах мыши, что позволяет предположить, что SMCHD1 способствует обогащению h4K27me3 во время XCI (Sakakibara et al., 2018; Гдула и др., 2019). В других гетерохроматических областях генома SMCHD1 локализуется совместно с репрессивной гистоновой меткой, h4K9me3, процесс, опосредованный LRIF1 (лиганд-зависимый фактор взаимодействия ядерных рецепторов 1). Однако SMCHD1 остается обогащенным по Xi, даже когда взаимодействия SMCHD1-LRIF1 нарушены, что указывает на альтернативный механизм, с помощью которого SMCHD1 нацелен на Xi, возможно, через убиквитинирование гистона h3AK119 (Brideau et al., 2015; Jansz et al., 2018) . Два более поздних события, которые блокируют подавление Xi, — это замена гистона h3A на макрогистон h3A и метилирование ДНК CpG-островков на DNMT3B (рис. 1A; Gartler and Riggs, 1983; Costanzi and Pehrson, 1998; Gendrel et al., 2012).
Dxz4
Во время установления XCI Xi конденсируется в двух супердоменах дальних контактов, разделенных областью, содержащей консервативный микросателлитный повтор Dxz4 lncRNA (Рисунок 1B; Deng et al., 2015; Minajigi et al., 2015; Darrow et al., 2016; Giorgetti et al., 2016). Эта двудольная конфигурация присутствует как у мышей, так и у человека, хотя и с разными размерами супердоменов, и делеция Dxz4 , специфически из Xi, приводит к нарушению двудольной структуры у обоих видов, что указывает на консервативную функцию (Darrow et al., 2016; Джорджетти и др., 2016; Bonora et al., 2018). Удаленный Xi приобретает конфигурацию, которая напоминает Xa с улучшенными TAD и отсеками, но только частично, что позволяет предположить, что, помимо Dxz4 , конфигурацией Xi управляют другие факторы.
CTCF-опосредованные взаимодействия между локусами Dxz4 / DXZ4 и другими X-сцепленными локусами, по-видимому, являются неотъемлемой частью формирования петель хроматина для упаковки Xi. Действительно, Dxz4 / DXZ4 связывают белок CTCF с цинковыми пальцами и компоненты кольцевого когезинового комплекса только на Xi (Horakova et al., 2012а, б). В другом месте генома конвергентные связывающие мотивы CTCF в основании петли хроматина явно способствуют сильным взаимодействиям, а инверсия сайтов CTCF нарушает образование петель (de Wit et al., 2015). Локус мыши Dxz4 содержит банк мотивов CTCF, расположенных в тандемной ориентации, тогда как локус человека содержит два банка мотивов с разной ориентацией (Horakova et al., 2012a, b). Наша группа сообщила, что инверсия мышиного локуса Dxz4 приводит к массивной инверсии дальних контактов, указывая на то, что локус Dxz4 сам по себе действует как структурная платформа для частых дальних контактов с множественными X-сцепленными локусами в направление продиктовано ориентацией его мотивов CTCF (Bonora et al., 2018). Закрепление петель хроматина размером с мегабазу на Dxz4 вызывает появление линии (или пламени), исходящей из Dxz4 на карте контактов (Рис. 1B). Остается определить, будут ли контакты между Dxz4 и др. X-сцепленными локусами быстро колебаться в индивидуальных клетках. Удивительно, но делеция Dxz4 вызывает лишь незначительную реактивацию X-сцепленных генов и незначительные изменения в экспрессии генов ускользания (Giorgetti et al., 2016; Bonora et al., 2018). В более позднем исследовании не сообщалось об изменениях в экспрессии генов вообще, что свидетельствует о несоответствии между клеточными линиями (Froberg et al., 2018). Более того, мыши с делецией Dxz4 на Xi не обнаруживают явного фенотипа (Andergassen et al., 2019). Таким образом, двудольная структура Xi не имеет четкой функции в регуляции генов Xi на данный момент. Однако сохранение локуса и двудольной структуры между человеком и мышью предполагает сохранение функции.
Фирре
Firre — еще один X-связанный локус lncRNA, который влияет на эпигенетические особенности и трехмерную структуру Xi.Мы и другие показали, что Firre транскрибируется только с активной Х-хромосомы (Calabrese et al., 2012; Froberg et al., 2018; Andergassen et al., 2019; Fang et al., 2019). Сообщалось о множественных изоформах транскриптов Firre , включая днРНК и кольцевые РНК (Izuogu et al., 2018). Как и Dxz4 , область Firre содержит множество локальных повторов, включая повтор R0, который привлекает организаторы хроматина CTCF и YY1, а также RAD21, компонент кольцевого комплекса когезина (Yang et al., 2015; Hacisuleyman et al., 2016). Локус Firre рекрутирует белок ядерного матрикса hnRNPU и взаимодействует со многими областями генома, что может объяснить, почему истощение РНК Firre вызывает широко распространенную дисрегуляцию аутосомных генов (Hacisuleyman et al., 2014; Andergassen et al., 2019; Fang et al. др., 2019; Левандовски и др., 2019). Что касается XCI, истощение РНК Firre в дифференцированных фибробластах не нарушает покрытие Xist или молчание гена; однако потеря h4K27me3 наблюдается на Xi (Yang et al., 2015; Fang et al., 2019). Напротив, эмбриональные стволовые клетки, лишенные Firre перед дифференцировкой, не обнаруживают изменений в h4K27me3 (Froberg et al., 2018). Вместе эти результаты подтверждают роль РНК Firre в поддержании, но не в инициации обогащения h4K27me3 на Xi в дифференцированных клетках.
Firre РНК также важна для поддержания положения Xi в ядре. Действительно, истощение Firre в дифференцированных клетках вызывает уменьшение перинуклеолярной и ядерной периферийной ассоциации Xi (Yang et al., 2015; Fang et al., 2019). Перинуклеолярный и периферийный компартменты часто связаны с гетерохроматином, а расположение ядрышек Xi, как предполагается, важно для точной репликации его эпигенетического состояния (Figure 1C; Zhang et al., 2007; de Wit and van Steensel, 2009). Ассоциация Xi с пластинкой может быть облегчена рецептором ламина B (LBR), рекрутируемым РНК Xist (Chen C.K. et al., 2016). Однако как активные, так и неактивные Х-хромосомы предпочтительно расположены вблизи периферии ядра, что позволяет предположить, что факторы, не связанные с XCI, могут контролировать позиционирование (Bischoff et al., 1993). Удивительно, но связывание одного или обоих аллелей XIC с ядерной пластиной через слитый белок TetR-EGFP-LaminB1 не нарушает инициацию XCI, что может означать, что спаривание XIC и посещение ядрышка не являются существенными для инициации XCI (Pollex and Heard , 2019).
Удаление Firre на Xi не нарушает двустороннюю организацию Xi, но вызывает локальные изменения в распределении контактов, которые могут отражать нарушение дальних контактов между Firre и Dxz4 , которые, как предполагается, обеспечивают уплотнение Xi (Дарроу и др., 2016; Barutcu et al., 2018; Bonora et al., 2018; Froberg et al., 2018; Fang et al., 2019). Контакты на большие расстояния между локусами могут помочь изолировать Xi в определенном компартменте или фазе ядра либо вблизи периферии ядра, либо в ядрышке. Локусы lncRNA Xist , Firre и Dxz4 могут играть согласованную роль в конденсации и изоляции Xi в определенной фазе, что д. Обеспечивать дифференциальную регуляцию двух Х-хромосом. Немногие исследования напрямую проверяли такую гипотезу из-за проблем с дизайном экспериментов (Heard et al., 2004).
Обнаружение генов, которые избегают инактивации X с помощью анализа отдельных клеток
Несмотря на то, что они локализованы в постоянно репрессированной среде, избранная подмножество генов развило механизмы, позволяющие избегать молчания и, таким образом, оставаться экспрессируемыми из Xi. Ряд генов избегает XCI индивидуальным, тканевым и клеточным типом, что может вызывать половые различия в экспрессии генов (Berletch et al., 2015; Cotton et al., 2015; Tukiainen et al., 2017 ; Balaton et al., 2018). У мышей 3-7% X-сцепленных генов избегают транскрипционного молчания, в то время как это число увеличивается до 20-30% у человека (Berletch et al., 2015; Balaton and Brown, 2016). Сравнение между видами показывает, что основной набор генов ускользает от XCI в большинстве клеток и тканей у млекопитающих, тогда как другие гены различаются между типами клеток, тканями и видами. Новые подходы, основанные на секвенировании одноклеточной РНК (scRNA-seq) в сочетании с анализом экспрессии аллельных генов SNP (однонуклеотидный полиморфизм), предоставляют данные о тысячах отдельных клеток различных типов и, таким образом, обещают более полную картину различий в статусе ускользания между типы клеток (рис. 2А).Присвоение статуса ускользания данному гену формально требует обнаружения двуаллельных чтений в отдельных клетках или в ткани с полностью искаженным XCI. Основное преимущество одноклеточных подходов состоит в том, что ткани со случайным XCI можно анализировать, но выпадение аллелей может вызвать проблемы для генов с низкой экспрессией (Reinius and Sandberg, 2015). Ограниченный анализ scRNA-seq около 1000 клеток, представляющих два типа клеток человека, выявил выявленные гены (например, FHL1 и ATP6AP2 ) с неполным XCI в подмножестве клеток, а также подтвердил гетерогенность XCI для TIMP1 (Tukiainen и другие., 2017). Другой аллельный scRNA-seq в человеческих фибробластах мало пересекается с другими исследованиями в отношении идентифицированных генов ускользания, что подчеркивает сложность достижения консенсуса (Wainer Katsir and Linial, 2019). Вариабельность ускользания от XCI у разных людей была подтверждена в анализе экспрессии аллелей одноклеточных у пяти человек (Garieri et al., 2018).
Рисунок 2. Распределение и структура генов ускользания. (A) Single-cell RNA-seq (scRNA-seq) позволяет определять закономерности ухода от XCI в типах клеток в ткани.Начиная с ткани, например мозга, отдельные типы клеток (окрашенные в розовый, светло-розовый и синий цвета) могут быть идентифицированы с помощью scRNA-seq. Последующий анализ SNP определяет статус ускользания каждого X-связанного гена на основе считываний из Xa и Xi (ген ускользания, esc, окрашен зеленым; инактивированный ген, неактивный, окрашен в черный цвет; ген, который частично ускользает, ∼esc, окрашен в светло-зеленый цвет. ). Каждый тип клеток может быть связан со статусом ускользания ансамбля X-сцепленных генов. Например, конкретный тип клеток (розовый) показывает выход из примера гена (обведен зеленым), а другой тип клеток (синий) показывает инактивацию того же гена (обведен черным).Комбинация данных об ансамбле генов потенциально может определять половые различия в конкретном типе клеток. (B) 3D-модель Xi с разрешением 1 МБ в мозге мыши, окрашенная для отображения плотности связывания аллельного CTCF (красный цвет указывает на большее связывание, а синий — меньшее связывание). Видны две области конденсации, разделенные расстоянием Dxz4 на шарнире. Белые точки обозначают концы хромосом, оранжевая точка — Dxz4 , а зеленые точки — гены ускользания. Гены ускользания, как правило, располагаются вне трехмерной структуры Xi [адаптировано из опубликованного рисунка (Deng et al., 2015) в Genome Biology под лицензией Springer Nature Publishing License: http://creativecommons.org/licenses/by/4.0/]. (C) Частичные карты контактов Hi-C (разрешение 4 Мб) активной (Ха) и неактивной (Xi) Х-хромосомы в области 4 Мб вокруг гена ускользания Ddx3x и инактивированного гена Eda2r подчеркивают ослабление TAD (синий) на Xi мыши, за исключением гена ускользания Ddx3x , где TAD видны как на Xi, так и на Xa. Цветовая шкала показывает нормализованное количество контактов (синий — большее количество контактов, красный — меньшее количество контактов) [адаптировано из рисунка (Bonora et al., 2018), опубликованной в Nature Communications под лицензией Springer Nature Publishing License: http://creativecommons.org/licenses/by/4.0/].
Исследования scRNA-seq развития отслеживали прогрессию XCI во время эмбриогенеза. Исследование доимплантационных эмбрионов человека показывает, что экспрессия биаллельного X-сцепленного гена может сохраняться до стадии бластоцисты, вместе с подавлением обоих аллелей (Petropoulos et al., 2016). Однако повторный анализ этих и дополнительных данных предоставил более подробную панораму случайных XCI от человеческого ооцита до бластоцисты, демонстрируя прогрессивное установление моноаллельной экспрессии гена, сцепленного с X, и позитивной регуляции X для поддержания баланса экспрессии по всему геному ( Морейра де Мелло и др., 2017; Чжоу и др., 2019). К сожалению, в эти исследования включено очень мало информации о генах ускользания, несмотря на выводы о значительных половых различиях в общей экспрессии генов. У мышей динамика XCI во время дифференцировки эмбриональных стволовых клеток обнаруживает постепенное снижение экспрессии генов ускользания, что согласуется с частичным распространением молчания (Chen G. et al., 2016).
Структурные и эпигенетические особенности генов ускользания
Ожидается, что гены, экспрессируемые из Xi, лишены эпигенетических сигнатур, характерных для инактивированных генов, и, по-видимому, расположены вдали от репрессивных геномных элементов.Недавнее исследование с использованием аллель-специфичного PRO-seq и прогнозирующего машинного обучения показывает, что основные детерминанты выхода из XCI включают расстояние от Xist и плотность элементов LINE (Sousa et al., 2019). Интересно, что гены ускользания часто группируются в домены, что является обычным явлением у человека, в то время как гены ускользания у мышей часто изолированы (Tsuchiya et al., 2004; Prothero et al., 2009; Berletch et al., 2011). Как правило, в ускользающих областях отсутствуют репрессивные гистоновые метки, такие как h4K27me3, и они обогащены метками активных гистонов, такими как ацетилирование, и метками элонгации транскрипции, включая РНК PolII S2P и h4K36me3 (Рисунок 1A; Disteche and Berletch, 2015; Sousa et al., 2019). Гипометилирование ДНК CpG-островков является надежным предиктором статуса ускользания, который успешно использовался для идентификации генов ускользания в массиве тканей человека, где аллельный анализ затруднен из-за небольшого числа SNP и / или отсутствия перекоса XCI (Cotton et al., 2015; Дункан и др., 2018). Удивительно, но гены ускользания в головном мозге и печени принимают специфические сигнатуры метилирования ДНК, которые включают обогащение не-CG гиперметилированием (mCH) по всему телу их гена, что может помочь поддерживать открытую структуру хроматина (Keown et al., 2017; Дункан и др., 2018).
гены ускользания, как правило, располагаются снаружи уплотненной инактивированной внутренней части Xi (рис. 2B; Chaumeil et al., 2006; Heard and Bickmore, 2007; Splinter et al., 2011; Deng et al., 2015; Bonora and Disteche, 2017). Однако еще предстоит определить, как именно трехмерная структура Xi влияет на склонность к побегу из XCI. Одним из факторов может быть повтор E Xist , который необходим для локализации ASh3L, компонента гистон-метилтрансферазы, который метилирует h4K4 для повышения экспрессии (Yue et al., 2017). Могут быть задействованы и другие факторы, например, lncRNAs, часто обнаруживаемые рядом с генами ускользания (Reinius et al., 2010). Ускользающие гены часто совместно локализуются с кластерами связывания CTCF и с TAD, что указывает на локальную компартментализацию Xi (рис. 2C; Giorgetti et al., 2016; Bonora et al., 2018). Однако делеция Dxz4 и потеря двудольной структуры Xi вызывает незначительное нарушение экспрессии гена ускользания или совсем не вызывает его (Giorgetti et al., 2016; Bonora et al., 2018; Froberg et al., 2018).
Становится все более очевидным, что внутренние генетические элементы ускользания действуют в cis , чтобы облегчить экспрессию из Xi (Balaton et al., 2018). Мы и другие предположили, что распространение сайленсинга в область ускользания или наоборот распространение активности гена на заглушенный ген может быть предотвращено элементами инсулятора, такими как CTCF или YY1, но этих элементов может быть недостаточно, и механизмы могут отличаться между генами ускользания. (Филиппова и др., 2005; Li, Carrel, 2008; Horvath et al., 2013; Chen C.Y. и др., 2016). Интересно, что ВАС, несущий человеческий ген ускользания RPS4X , вставленный в заглушенный локус Hprt у мыши, сохраняет статус ускользания in vivo как в течение начала XCI, так и в течение жизни мыши (Peeters et al., 2018) . Будет интересно идентифицировать очевидно консервативные инсуляторные элементы, участвующие в этом процессе, и проверить их роль в формировании областей ускользания, например, путем выделения их в отдельные петли хроматина или фазы в ядре.
Роль генов, связанных с Х-хромосомой, в различиях полов и в заболеваниях
Одним из основных последствий бегства от XCI является дифференциальная экспрессия генов у мужчин и женщин (Mele et al., 2015). Недавнее исследование, основанное на тысячах транскриптомов, охватывающих 29 тканей человека, дает подробный обзор экспрессии генов с предвзятым отношением к полу у людей и демонстрирует, что экспрессия генов ускользания обычно смещена в сторону женщин (Tukiainen et al., 2017). Однако подмножество генов ускользания, расположенных в псевдоавтосомной области, разделяемой между X- и Y-хромосомами, смещено в сторону мужчин, вероятно, из-за более низкой экспрессии у женщин из-за распространения сайленсинга на Xi (Tukiainen et al., 2017). Биологические последствия этого предубеждения по признаку пола остаются в значительной степени неизученными. Хотя очевидно, что определенные гены ускользания, например Kdm6a и его Y-паралог Uty , экспрессируются в разных частях мозга мышей, их роль в фенотипических половых различиях не выяснена (Xu et al., 2008). ). Фактически, несколько признаков были связаны с половым уклоном в экспрессии X-сцепленного гена у нормальных здоровых людей. Одним из примеров является долголетие, недавние данные свидетельствуют о том, что наличие двух Х-хромосом продлевает продолжительность жизни независимо от пола гонад.Это было продемонстрировано с использованием четырехъядерного генотипа (FCG), модели мышей, способной дифференцировать эффекты гормонов по сравнению с комплементом половых хромосом, который показал, что мыши XX с яичниками или семенниками жили дольше, чем мыши XY любого гонадного фенотипа (Davis et al. др., 2019). Кроме того, наличие двух Х-хромосом приводит к улучшению регуляции артериального давления и увеличению способности ослаблять последствия травм головного мозга (Pessoa et al., 2015; McCullough et al., 2016).
Что касается восприимчивости к болезням, имеется достаточно доказательств того, что предвзятость пола в экспрессии X-сцепленного гена играет роль.Например, наличие двух Х-хромосом увеличивает риск развития аутоиммунитета [см. Обзор Syrett and Anguera (2019)]. Этот феномен может быть результатом необычных паттернов XCI в иммунных клетках, которые могут оставлять определенные X-сцепленные гены, участвующие в иммунном ответе, чувствительными к реактивации (Wang et al., 2016; Syrett et al., 2019). Это подтверждается исследованиями, демонстрирующими, что ген Toll-подобного рецептора 7, который ускользает от XCI в лимфоцитах человека, вызывает системную красную волчанку при сверхэкспрессии в моделях мышей (Deane et al., 2007; Souyris et al., 2018). Х-хромосома несет в себе большое количество генов, важных для функции мозга, а дозировка некоторых генов ускользания влияет на неврологические фенотипы, такие как судороги и расстройство аутистического спектра (Shoubridge et al., 2019). Например, мутации потери функции в гене ускользания, IQSEC2 , способствуют проявлению фенотипов, которые включают умеренную и тяжелую умственную отсталость (Shoubridge et al., 2019). Другой пример — KDM5C , ген ускользания, который кодирует гистоновую деметилазу и регулирует развитие и функцию нейронов (Iwase et al., 2016; Scandaglia et al., 2017; Ким и др., 2018). Мутации в KDM5C вызывают умственную отсталость у мужчин и женщин, что свидетельствует о дозовой чувствительности этого гена (Brookes et al., 2015).
Вариабельность экспрессии генов ускользания может способствовать половым различиям в предрасположенности к определенным видам рака (Arnold and Disteche, 2018). Многие типы рака имеют тенденцию к половому признаку по своей природе, а некоторые из них могут быть объяснены мутациями в X-сцепленных генах ускользания, называемых EXITS (Escape from X Inactivation Tumor Suppressor) (Таблица 1; Dunford et al., 2017). Одним из примеров является рак мочевого пузыря, заболеваемость которым у мужчин в три-пять раз выше, чем у женщин (Edgren et al., 2012). Недавние исследования на мышах FCG показывают, что мыши XX с раком мочевого пузыря выживают с большей скоростью по сравнению с мышами XY независимо от пола гонад, предполагая, что дозировка X-сцепленного гена является внутренним детерминантом выживания (Kaneko and Li, 2018). Интересно, что женский ген ускользания KDM6A является сильным опухолевым супрессором, который действует через зависимые от деметилирования и независимые механизмы, снижая пролиферацию клеток рака мочевого пузыря (Kaneko and Li, 2018).
Таблица 1. Гены ускользания, не относящиеся к PAR, как опухолевые супрессоры при мужском раке.
Нарушения числа половых хромосом, такие как синдром Тернера (45, X) и синдром Клайнфельтера (47, XXY), напрямую связаны с генами ускользания в аномальных фенотипах, поскольку в этих условиях такие гены будут гапло-недостаточными и сверхэкспрессируемыми, соответственно. Одной из отличительных черт синдрома Тернера является преждевременное нарушение функции яичников и бесплодие. Х-хромосома в три раза более обогащена генами, экспрессируемыми в женских репродуктивных органах, по сравнению с аутосомами, что указывает на ее роль в женской фертильности (Liu, 2019).Определение вклада конкретных X-сцепленных генов в аномальные фенотипы Тернера находится в стадии разработки. Например, гены ускользания KDM6A и TIMP1 предположительно участвуют в преждевременной недостаточности яичников и формировании аневризмы аорты, соответственно (Trolle et al., 2016; Viuff et al., 2019). Недавние исследования вариации числа копий, сцепленных с Х-хромосомой (CNV) в когортах здоровых женщин и женщин с первичной недостаточностью яичников (POI), показывают высокую распространенность делеций, охватывающих гены ускользания, а также днРНК (Яценко и др., 2019). Конечно, гены ускользания не будут единственными генами, сцепленными с Х, участвующими в бесплодии Тернера, поскольку все гены, сцепленные с Х, реактивируются в первичных половых клетках женщин, процесс, опосредованный PR-доменом, содержащим белок 14 (PRDM14) для удаления h4K27me3 на Си (Mallol et al., 2019). Бесплодие, наблюдаемое у людей Клайнфельтера, потенциально может быть результатом аномальной дозировки гена ускользания, но другим важным фактором может быть аномальное мейотическое спаривание половых хромосом. Интересно, что любые аномальные числа Х-хромосом (XXY, XXX или X) вызывают общее нарушение паттернов метилирования ДНК в аутосомных генах, демонстрируя широко распространенные эпигенетические эффекты X-анеуплоидии (Trolle et al., 2016; Skakkebaek et al., 2018). Кроме того, количество X- и / или Y-хромосом влияет на пространственную конформацию хромосомы, особенно активной X-хромосомы, но роль этого структурного изменения не выяснена (Jowhar et al., 2018). В совокупности эти данные указывают на то, что неправильная дозировка X-сцепленного гена является причинным фактором фенотипов заболевания, начиная от функции мозга, предрасположенности к раку и заканчивая нарушением фертильности.
Перспектива
Хотя многое было изучено о различных lncRNAs и белках, которые контролируют структурные и эпигенетические особенности Xi и его молчание, их точные способы действия еще предстоит изучить.С точки зрения трехмерной структуры Xi будет очень интересно определить факторы, участвующие в ядерной компартментализации и разделении фаз. Необходимы дополнительные эксперименты, чтобы связать конкретную трехмерную структуру и расположение ядра Xi с его отличным эпигенетическим ландшафтом. Более того, мало что известно о контактах каждой Х-хромосомы с остальной частью генома в женских клетках и тканях, а также о таких контактах с гетерохроматической Y-хромосомой в мужских клетках и тканях.Тканевые различия в расположении хромосом в ядре плохо изучены, и лишь несколько функциональных исследований изучали последствия манипулирования положением хромосом. Эпигенетический контроль бегства от XCI также требует дальнейших функциональных исследований. К сожалению, текущие исследования по идентификации генов ускользания в определенных типах клеток и тканях ограничены из-за относительно небольшого количества информативных полиморфизмов у человека. Однако одноклеточные анализы развиваются быстрыми темпами, и некоторые методы позволяют проводить анализ тысяч клеток в тканях для создания карт экспрессии или доступности генов во всем организме, как показано, например, в недавно опубликованном атласе тканей мышей / типы клеток (Cusanovich et al., 2018). Хотя есть убедительные доказательства половых различий в восприимчивости к болезням, понимание роли отдельных сцепленных с полом генов потребует тщательного изменения их дозировки.
Авторские взносы
JB, HF и CD обрисовали в общих чертах и написали обзор. Все авторы одобрили окончательную рукопись.
Финансирование
Авторство этого обзора поддержано грантами GM131745 от Национальных институтов общей медицины и DK107979 от 4D Nucleome Program Общего фонда национальных институтов здравоохранения.
Конфликт интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим Галу Филиппову, Синьсянь Дэн и Ди Ким Нгуен за критическое прочтение рукописи и конструктивные редакционные комментарии. Мы также благодарим Wenxiu Ma и Giancarlo Bonora за первоначальное создание карт контактов Hi-C и трехмерной модели фигур (см. Ссылки в подписях к рисункам).
Список литературы
Андергассен, Д., Смит, З., Левандовски, Дж., Герхардингер, К., Мейснер, А., и Ринн, Дж. (2019). In vivo firre и делеция Dxz4 проясняет роль регуляции аутосомных генов. bioRxiv DOI: 10.1101 / 612440
CrossRef Полный текст | Google Scholar
Баруцу, А. Р., Маасс, П. Г., Левандовски, Дж. П., Вайнер, К. Л., и Ринн, Дж. Л. (2018). Граница TAD сохраняется при делеции богатого CTCF елового локуса. Nat.Commun. 9: 1444. DOI: 10.1038 / s41467-018-03614-3610
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берлетч, Дж. Б., Ма, В., Янг, Ф., Шендур, Дж., Ноубл, В. С., Дистеч, К. М. и др. (2015). Спасение от инактивации x варьируется в тканях мышей. PLoS Genet. 11: e1005079. DOI: 10.1371 / journal.pgen.1005079
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берлетч, Дж. Б., Янг, Ф., Сюй, Дж., Каррел, Л., и Дистеч, К.М. (2011). Гены, которые избегают инактивации X. Хум. Genet. 130, 237–245. DOI: 10.1007 / s00439-011-1011-z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бишофф А., Альберс Дж., Харбоуш И., Стельцер Э., Кремер Т. и Кремер К. (1993). Различия в размерах и форме активных и неактивных доменов Х-хромосомы в ядрах клеток амниотической жидкости человека. Microsc. Res. Tech. 25, 68–77. DOI: 10.1002 / jemt.1070250110
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Блевитт, М.Е., Гендрел, А. В., Панг, З., Воробей, Д. Б., Уайтлоу, Н., Крейг, Дж. М. и др. (2008). SmcHD1, содержащий шарнирный домен структурного поддержания хромосом, играет критическую роль в инактивации X. Nat Genet. 40, 663–669. DOI: 10,1038 / нг.142
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бонора, Г., Дэн, X., Фанг, Х., Рамани, В., Цю, Р., Берлетч, Дж. Б. и др. (2018). Зависимые от ориентации контакты Dxz4 формируют трехмерную структуру неактивной Х-хромосомы. Nat. Commun. 9: 1445. DOI: 10.1038 / s41467-018-03694-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Борсани Г., Тонлорензи Р., Симмлер М. К., Дандоло Л., Арно Д., Капра В. и др. (1991). Характеристика мышиного гена, экспрессируемого неактивной Х-хромосомой. Природа 351, 325–329. DOI: 10.1038 / 351325a0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бридо, Н. Дж., Кокер, Х., Гендрел, А.V., Siebert, C.A., Bezstarosti, K., Demmers, J., et al. (2015). Независимые механизмы направляют SMCHD1 на триметилированный гистон h4, модифицированный лизином 9 хроматин и неактивную Х-хромосому. Мол. Cell Biol. 35, 4053–4068. DOI: 10.1128 / MCB.00432-415
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брокдорф, Н., Эшворт, А., Кей, Г. Ф., Купер, П., Смит, С., МакКейб, В. М. и др. (1991). Сохранение положения и исключительная экспрессия xist мыши из неактивной Х-хромосомы. Природа 351, 329–331. DOI: 10.1038 / 351329a0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Brookes, E., Laurent, B., Ounap, K., Carroll, R., Moeschler, J. B., Field, M., et al. (2015). Мутации в гене умственной отсталости KDM5C снижают стабильность белка и активность деметилазы. Хум. Мол. Genet. 24, 2861–2872. DOI: 10.1093 / hmg / ddv046
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Браун, К.Дж., Лафренье, Р. Г., Пауэрс, В. Э., Себастио, Г., Баллабио, А., Петтигрю, А. Л. и др. (1991). Локализация центра инактивации X на X-хромосоме человека в Xq13. Природа 349, 82–84. DOI: 10.1038 / 349082a0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Калабрезе, Дж. М., Сан, У., Сонг, Л., Магфорд, Дж. У., Уильямс, Л., Йи, Д. и др. (2012). Сайт-специфическое молчание регуляторных элементов как механизм инактивации X. Ячейка 151, 951–963.DOI: 10.1016 / j.cell.2012.10.037
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Церасе, А., Армаос, А., Ноймайер, К., Авнер, П., Гуттман, М., и Тарталья, Г. Г. (2019). Разделение фаз приводит к инактивации Х-хромосомы: гипотеза. Nat. Struct. Мол. Биол. 26, 331–334. DOI: 10.1038 / s41594-019-0223-220
CrossRef Полный текст | Google Scholar
Шомей Дж., Ле Баккон П., Вутц А. и Херд Э. (2006). Новая роль РНК Xist в формировании репрессивного ядерного компартмента, в который гены рекрутируются при замалчивании. Genes Dev. 20, 2223–2237. DOI: 10.1101 / gad.380906
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чен, К. К., Бланко, М., Джексон, К., Азнаурян, Э., Олликайнен, Н., Сурка, К. и др. (2016). Xist рекрутирует Х-хромосому в ядерную пластинку, чтобы обеспечить молчание по всей хромосоме. Наука 354, 468–472. DOI: 10.1126 / science.aae0047
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чен, К. Ю., Ши, В., Балатон, Б. П., Мэтьюз, А. М., Ли, Ю., Аренильяс, Д. Дж. И др. (2016). Связь связывания YY1 со смещенной по полу транскрипцией выявляется с помощью уровней X-сцепленного транскрипта и анализа аллельного связывания. Sci. Реп. 6: 37324. DOI: 10.1038 / srep37324
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чен, Г., Шелл, Дж. П., Бенитес, Дж. А., Петропулос, С., Йилмаз, М., Рейниус, Б. и др. (2016). Одноклеточный анализ динамики инактивации Х-хромосомы и плюрипотентности во время дифференцировки. Genome Res. 26, 1342–1354. DOI: 10.1101 / gr.201954.115
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чу, К., Чжан, К. К., да Роча, С. Т., Флинн, Р. А., Бхарадвадж, М., Калабрезе, Дж. М. и др. (2015). Систематическое открытие белков, связывающих РНК Xist. Ячейка 161, 404–416. DOI: 10.1016 / j.cell.2015.03.025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Клемсон, К. М., Холл, Л. Л., Байрон, М., МакНил, Дж.и Лоуренс Дж. Б. (2006). Х-хромосома организована в виде богатого генами внешнего обода и внутреннего ядра, содержащих заглушенные негенные последовательности. Proc. Natl. Акад. Sci. США 103, 7688–7693. DOI: 10.1073 / pnas.0601069103
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Колонори Д., Суну Х., Криз А. Дж., Ван С. Ю. и Ли Дж. Т. (2019). Делеционный анализ Xist выявляет взаимозависимость между xist РНК и комплексами polycomb в распространении вдоль неактивного X. Мол. Ячейка 74, 101.E – 117.E. DOI: 10.1016 / j.molcel.2019.01.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коттон, А. М., Ге, Б., Лайт, Н., Адуэ, В., Пастинен, Т., и Браун, К. Дж. (2013). Анализ экспрессируемых SNP позволяет идентифицировать различные степени экспрессии неактивной Х-хромосомы человека. Genome Biol. 14: R122. DOI: 10.1186 / GB-2013-14-11-r122
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хлопок, А.М., Прайс, Э. М., Джонс, М. Дж., Балатон, Б. П., Кобор, М. С., Браун, К. Дж. (2015). Пейзаж метилирования ДНК на Х-хромосоме отражает плотность CpG, функциональное состояние хроматина и инактивацию Х-хромосомы. Хум. Мол. Genet. 24, 1528–1539. DOI: 10.1093 / hmg / ddu564
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кусанович, Д. А., Хилл, А. Дж., Агамирзайе, Д., Даза, Р. М., Плинер, Х. А., Берлетч, Дж. Б. и др. (2018). Одноклеточный атлас доступности хроматина млекопитающих in vivo. Ячейка 174, 1309–1324.e18. DOI: 10.1016 / j.cell.2018.06.052
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дэрроу, Э. М., Хантли, М. Х., Дудченко, О., Стаменова, Э. К., Дюран, Н. К., Сан, З. и др. (2016). Делеция DXZ4 на неактивной Х-хромосоме человека изменяет архитектуру генома более высокого порядка. Proc. Natl. Акад. Sci. США 113, E4504 – E4512. DOI: 10.1073 / pnas.1609643113
PubMed Аннотация | CrossRef Полный текст | Google Scholar
де Вит, Э., и ван Стинзель, Б. (2009). Домены хроматина у высших эукариот: выводы из исследований картирования всего генома. Хромосома 118, 25–36. DOI: 10.1007 / s00412-008-0186-180
PubMed Аннотация | CrossRef Полный текст | Google Scholar
de Wit, E., Vos, E. S., Holwerda, S. J., Valdes-Quezada, C., Verstegen, M.J., Teunissen, H., et al. (2015). Полярность связывания CTCF определяет образование петель хроматина. Мол. Ячейка 60, 676–684. DOI: 10.1016 / j.molcel.2015.09.023
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дин, Дж.A., Pisitkun, P., Barrett, R. S., Feigenbaum, L., Town, T., Ward, J. M., et al. (2007). Контроль экспрессии толл-подобного рецептора 7 необходим для ограничения аутоиммунитета и пролиферации дендритных клеток. Иммунитет 27, 801–810. DOI: 10.1016 / j.immuni.2007.09.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дэн, X., Берлетч, Дж. Б., Нгуен, Д. К., и Дистеч, К. М. (2014). Регуляция Х-хромосомы: различные паттерны в развитии, тканях и заболеваниях. Nat. Преподобный Жене. 15, 367–378. DOI: 10.1038 / nrg3687
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дэн, X., Ма, В., Рамани, В., Хилл, А., Янг, Ф., Ай, Ф. и др. (2015). Двудольная структура неактивной X-хромосомы мыши. Genome Biol. 16: 152. DOI: 10.1186 / s13059-015-0728-728
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Диксон, Дж. Р., Сельварадж, С., Юэ, Ф., Ким, А., Ли, Ю., Шен, Ю., и др.(2012). Топологические домены в геномах млекопитающих, идентифицированные путем анализа взаимодействий хроматина. Природа 485, 376–380. DOI: 10.1038 / nature11082
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дункан, К. Г., Гримм, С. А., Морган, Д. Л., Бушел, П. Р., Беннет, Б. Д., Программа, Н. С. С. и др. (2018). Компенсация дозировки и ландшафт метилирования ДНК Х-хромосомы в печени мыши. Sci. Отчет 8: 10138. DOI: 10.1038 / s41598-018-28356-28353
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Данфорд, А., Вайншток, Д. М., Савова, В., Шумахер, С. Е., Клири, Дж. П., Йода, А. и др. (2017). Гены-супрессоры опухолей, которые избегают X-инактивации, вносят свой вклад в предвзятость пола при раке. Nat. Genet. 49, 10–16. DOI: 10,1038 / нг.3726
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Энгрейц, Дж. М., Олликайнен, Н., Гутман, М. (2016). Длинные некодирующие РНК: пространственные усилители, контролирующие структуру ядра и экспрессию генов. Nat. Преподобный Мол. Cell Biol. 17, 756–770. DOI: 10.1038 / nrm.2016.126
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фанг, Х., Бонора, Г., Левандовски, Дж., Такур, Дж., Филиппова, Г. Н., Хеникофф, С., и др. (2019). Транс- и цис-действующие эффекты днРНК усиливают эпигенетические и структурные особенности неактивной Х-хромосомы. bioRxiv DOI: 10.1101 / 687236
CrossRef Полный текст | Google Scholar
Филиппова Г. Н., Ченг, М. К., Мур, Дж. М., Чыонг, Дж.П., Ху, Ю. Дж., Нгуен, Д. К. и др. (2005). Границы между хромосомными доменами инактивации и ускользания X связывают CTCF и не имеют метилирования CpG во время раннего развития. Dev. Cell 8, 31–42.
PubMed Аннотация | Google Scholar
Фроберг, Дж. Э., Пинтер, С. Ф., Криз, А. Дж., Джегу, Т., и Ли, Дж. Т. (2018). Мегадомены и суперцепи формируются динамически, но не необходимы для инактивации Х-хромосомы и ускользания генов. Nat. Commun. 9: 5004. DOI: 10.1038 / s41467-018-07446-w
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бактериальные клетки разного размера, формы и расположения
Бактерии — это прокариотические одноклеточные микроорганизмы, в которых отсутствуют пигменты хлорофилла. Структура клетки проще, чем у других организмов, поскольку в ней нет ядер или мембраносвязанных органелл.
Благодаря наличию жесткой клеточной стенки бактерии сохраняют определенную форму, хотя они различаются по форме, размеру и структуре.
При просмотре под световым микроскопом большинство бактерий проявляются в трех основных формах: стержень (палочка), сфера (кокк) и спиральный тип (вибрион). Фактически, структура бактерий имеет два аспекта: расположение и форму. Что касается аранжировки, это могут быть парные (дипло), гроздья, похожие на виноград (стафило) или цепочки (стрепто). По форме они могут быть в основном палочками (бациллы), сферами (кокки) и спиралями (спириллум).
Размер бактериальной клетки
Средний диаметр сферических бактерий равен 0.5-2,0 мкм. Для палочковидных или нитчатых бактерий длина составляет 1–10 мкм, а диаметр — 0,25–1,0 мкм.
- E. coli , бацилла среднего размера составляет от 1,1 до 1,5 мкм в ширину и от 2,0 до 6,0 мкм в длину.
- Спирохеты иногда достигают 500 мкм в длину, а диаметр цианобактерии
- Oscillatoria составляет около 7 мкм.
- Бактерия, Epulosiscium fishelsoni , видна невооруженным глазом (длина 600 мкм, диаметр 80 мкм).
- У одной группы бактерий, называемых микоплазмами, есть особи, размер которых намного меньше этих размеров. Их размер составляет около 0,25 мкм, и они являются самыми маленькими ячейками из известных на сегодняшний день. Ранее они были известны как организмы, подобные плевропневмонии (PPLO).
- Mycoplasma gallicepticum, w размером примерно от 200 до 300 нм считаются самыми маленькими бактериями в мире.
- Thiomargarita namibiensis — крупнейшая в мире бактерия, грамотрицательная протеобактерия, обнаруженная в океанических отложениях у побережья Намибии.Обычно он составляет 0,1–0,3 мм (100–300 мкм) в поперечнике, но наблюдались более крупные клетки до 0,75 мм (750 мкм).
Таким образом, некоторые бактерии намного крупнее средней эукариотической клетки (типичные клетки растений и животных имеют диаметр от 10 до 50 мкм).
Форма бактериальной клетки
Три основных бактериальных формы — это кокковая (сферическая), палочкообразная (палочковидная) и спиральная (закрученная), однако плеоморфные бактерии могут принимать несколько форм.
Форма бактериальной клетки- Кокки (или кокк для отдельной клетки) представляют собой круглые клетки, иногда слегка сплющенные, когда они соседствуют друг с другом.
- Бациллы (или бациллы для одной клетки) представляют собой палочковидные бактерии.
- Спирилла (или спириллум для отдельной клетки) — изогнутые бактерии, которые могут варьироваться от слегка изогнутой формы до спиралевидной, похожей на штопор. Многие спириллы неподвижны и способны двигаться. Особая группа спирилл, известная как спирохеты, длинные, тонкие и гибкие.
Расположение кокков
Кокковые бактерии могут существовать поодиночке, парами (как диплококки), группами по четыре (как тетрады), цепочками (как стрептококки), группами (как стапилококки) или в кубиках, состоящих из восьми клетки (как сарцины).Кокки могут быть овальными, удлиненными или уплощенными с одной стороны. Кокки могут оставаться прикрепленными после деления клеток. Эти групповые характеристики часто используются для идентификации определенных кокков.
1. Диплококки
Кокки расположены попарно.
Примеры: Streptococcus pneumoniae, Moraxella catarrhalis , Neisseria gonorrhoeae, и т. Д.
2. Streptococci
Кокки расположены в цепочки, поскольку клетки делятся в одной плоскости.
Примеры: Streptococcus pyogenes, Streptococcus agalactiae
3. Tetrads
Кокки располагаются в пакетах по четыре клетки, так как клетки делятся на две равнины.
Примеры: Aerococcus, Pediococcus и Tetragenococcus
4. Sarcinae
Кокки расположены кубовидно, поскольку клетки образуются путем регулярных делений клеток в трех плоскостях. Кокки, которые делятся в трех плоскостях и остаются группами, напоминают группы по восемь кубиков.
Примеры: Sarcina ventriculi, Sarcina ureae и т. Д.
5. Стафилококки
Кокки расположены в виде гроздей винограда, образованных нерегулярными делениями клеток на трех равнинах.
Примеры: Staphylococcus aureus
Расположение бацилл
Цилиндрические или палочковидные бактерии называются «бациллами» (множественное число: бациллы).
1. Diplobacilli
24.1B: Структура и функции клеток грибов
Грибы представляют собой одноклеточные или многоклеточные гетеротрофные разлагатели с толстыми клетками, которые поедают разлагающееся вещество и образуют клубки нитей.
Задачи обучения
- Опишите физические структуры, связанные с грибами
Ключевые моменты
- Стенки клеток грибов жесткие и содержат сложные полисахариды, называемые хитином (придает структурной прочности) и глюканами.
- Эргостерол — это стероидная молекула в клеточных мембранах, которая заменяет холестерин, обнаруженный в мембранах клеток животных.
- Грибы могут быть одноклеточными, многоклеточными или диморфными, что означает, что грибы являются одноклеточными или многоклеточными в зависимости от условий окружающей среды.
- Грибы на морфологической вегетативной стадии состоят из клубка тонких нитевидных гиф, тогда как репродуктивная стадия обычно более очевидна.
- Грибы любят находиться во влажной и слабокислой среде; они могут расти как на свете, так и без кислорода.
- Грибы являются сапрофитными гетеротрофами в том смысле, что они используют мертвое или разлагающееся органическое вещество в качестве источника углерода.
Ключевые термины
- глюкан : любой полисахарид, представляющий собой полимер глюкозы
- эргостерол : функциональный эквивалент холестерина, обнаруженного в клеточных мембранах грибов и некоторых простейших, а также стероидный предшественник витамина D2
- мицелий : вегетативная часть любого гриба, состоящая из массы ветвящихся нитевидных гиф, часто подземных
- гиф : длинная ветвистая нитчатая структура гриба, являющаяся основным способом вегетативного роста
- перегородка : деление клеточной стенки между гифами гриба
- слоевище : вегетативное тело гриба
- сапрофит : любой организм, который живет за счет мертвого органического вещества, например, некоторые грибы и бактерии
- хитин : сложный полисахарид, полимер N-ацетилглюкозамина, обнаруженный в экзоскелете членистоногих и в клеточных стенках грибов; считается ответственным за некоторые формы астмы у людей
Структура и функции ячеек
Грибы — эукариоты со сложной клеточной организацией.Как и эукариоты, клетки грибов содержат связанное с мембраной ядро, в котором ДНК обернута вокруг гистоновых белков. Некоторые типы грибов имеют структуру, сопоставимую с бактериальными плазмидами (петли ДНК). Клетки грибов также содержат митохондрии и сложную систему внутренних мембран, включая эндоплазматический ретикулум и аппарат Гольджи.
В отличие от клеток растений, клетки грибов не содержат хлоропластов или хлорофилла. Многие грибы демонстрируют яркие цвета, обусловленные другими клеточными пигментами, от красного до зеленого и черного.Ядовитую Amanita muscaria (мухомор) можно узнать по ярко-красной шляпке с белыми пятнами. Пигменты у грибов связаны с клеточной стенкой. Они играют защитную роль от ультрафиолетового излучения и могут быть токсичными.
Рисунок \ (\ PageIndex {1} \): Ядовитая Amanita muscaria : Ядовитая Amanita muscaria произрастает в умеренных и северных регионах Северной Америки.Жесткие слои клеточных стенок грибов содержат сложные полисахариды, называемые хитином и глюканами.Хитин, также содержащийся в экзоскелете насекомых, придает структурную прочность клеточным стенкам грибов. Стенка защищает клетку от высыхания и хищников. У грибов есть плазматические мембраны, подобные другим эукариотам, за исключением того, что структура стабилизируется эргостеролом: молекулой стероида, которая заменяет холестерин, обнаруженный в мембранах клеток животных. Большинство членов королевства грибов неподвижны.
Рост
Вегетативное тело гриба представляет собой одноклеточное или многоклеточное слоевище.Диморфные грибы могут переходить из одноклеточного в многоклеточное состояние в зависимости от условий окружающей среды. Одноклеточные грибы обычно называют дрожжами. Saccharomyces cerevisiae (пекарские дрожжи) и Candida видов (возбудители молочницы, распространенной грибковой инфекции) являются примерами одноклеточных грибов.
Рисунок \ (\ PageIndex {1} \): Пример одноклеточного грибка : Candida albicans является дрожжевой клеткой и возбудителем кандидоза и молочницы.Этот организм имеет морфологию, сходную с кокковыми бактериями; однако дрожжи — это эукариотический организм (обратите внимание на ядро).Большинство грибов — многоклеточные организмы. У них есть две отчетливые морфологические стадии: вегетативная и репродуктивная. Вегетативная стадия состоит из клубка тонких нитевидных структур, называемых гифами (единичные, гифы), тогда как репродуктивная стадия может быть более заметной. Масса гиф — мицелий. Он может расти на поверхности, в почве или разлагающемся материале, в жидкости или даже на живой ткани.Хотя отдельные гифы необходимо наблюдать под микроскопом, мицелий гриба может быть очень большим, и некоторые виды действительно являются «огромным грибом». Гигантский Armillaria solidipes (опята) считается крупнейшим организмом на Земле, распространяющимся на более чем 2 000 акров подземной почвы в восточном Орегоне; по оценкам, ему не менее 2400 лет.
Рисунок \ (\ PageIndex {1} \): Пример мицелия гриба : Мицелий гриба Neotestudina rosati может быть патогенным для человека.Грибок проникает через порез или царапину и развивает мицетому, хроническую подкожную инфекцию.Большинство гиф грибов разделены на отдельные клетки торцевыми стенками, называемыми перегородками (единичные, перегородки) (а, в). У большинства типов грибов крошечные отверстия в перегородках обеспечивают быстрый поток питательных веществ и небольших молекул от клетки к клетке вдоль гифы. Их описывают как перфорированные перегородки. Гифы в формах для хлеба (принадлежащих к Phylum Zygomycota) не разделены перегородками. Вместо этого они образованы крупными клетками, содержащими множество ядер, расположение которых описано как гифы ценоцитов (b).Грибы процветают во влажной и слегка кислой среде; они могут расти как со светом, так и без него.
Рисунок \ (\ PageIndex {1} \): Разделение гиф на отдельные клетки : Гифы грибов могут быть (а) перегородками или (б) ценоцитарными (coeno- = «общий»; -cytic = «клеточный») с множеством ядра присутствуют в единственной гифе. Микрофотография в светлом поле (c) Phialophora richardsiae показывает перегородки, разделяющие гифы.Питание
Как и животные, грибы являются гетеротрофами: они используют сложные органические соединения в качестве источника углерода, а не фиксируют углекислый газ из атмосферы, как это делают некоторые бактерии и большинство растений.Кроме того, грибы не фиксируют азот из атмосферы. Как и животные, они должны получать его из своего рациона. Однако, в отличие от большинства животных, которые глотают пищу, а затем переваривают ее в специализированных органах, грибы выполняют эти шаги в обратном порядке: пищеварение предшествует приему пищи. Во-первых, экзоферменты выводятся из гиф, где они перерабатывают питательные вещества в окружающей среде. Затем более мелкие молекулы, образующиеся в результате этого внешнего переваривания, абсорбируются через большую площадь поверхности мицелия.Как и в случае с клетками животных, запасающий полисахарид — это гликоген, а не крахмал, содержащийся в растениях.
Грибы — это в основном сапробионты (сапрофит — эквивалентный термин): организмы, которые получают питательные вещества из разлагающихся органических веществ. Они получают свои питательные вещества из мертвых или разлагающихся органических веществ, в основном из растительного материала. Экзоферменты грибов способны расщеплять нерастворимые полисахариды, такие как целлюлоза и лигнин мертвой древесины, на легко усваиваемые молекулы глюкозы. Таким образом, углерод, азот и другие элементы попадают в окружающую среду.Из-за разнообразия метаболических путей грибы играют важную экологическую роль и изучаются как потенциальные инструменты для биоремедиации.
Некоторые грибы паразитируют, поражая растения или животных. Головня и голландский вяз поражают растения, тогда как микоз и кандидоз (молочница) являются важными с медицинской точки зрения грибковыми инфекциями у людей.
.