Как и чем размножаются папоротники в природе и домашних условиях: способы
Папоротники — это растения, которые живут на Земле сотни миллионов лет. Они за это время практически не изменили свое строение, только уменьшились в росте. На их примере люди могут увидеть, как формировалась жизнь на планете. Эти растения не смогли развить в себе способность цвести и плодоносить, и никому никогда не удастся найти распустившийся цветок папоротника.
Что такое папоротник
Наряду с обычным для растений органом корневищем у папоротниковых (Polypodióphyta) вместо стеблей, листьев, цветоносов с семенными коробочками развился листоподобный побег, получивший научное название вайя. На вайях образуются споры, с помощью которых происходит один из способов размножения и развития папоротников.
Отпечаток папоротника на камне
Сейчас на Земле идентифицировано около 10 тыс. многолетних видов папоротников. В умеренном климате России они растут во влажных тенистых лесах, их выращивают как садовую культуру.
К сведению! Выращивание орляка в условиях сада может негативно отразиться на домашних животных. В зеленых частях растения содержатся опасные для них ядовитые вещества.
Тропические виды Polypodióphyta отличаются большим разнообразием вай, могут иметь древовидные формы. Некоторые из них растут высоко над землей на ветвях других растений. В холодном климате выращивание и размножение теплолюбивых папоротников и папоротниковидных растений (хвоща, плауна) проводят в домашних условиях.
Среди популярных комнатных папоротников выделяются Аспелиум с побегами, напоминающими зеленый фонтан; Блехнум, вайи которого растут как листья у пальмы; Нефролепис — любитель большого свободного пространства; Дискония — высокорослое растение для офисных помещений; Платицериум, вайи которого подобны оленьим рогам.
Тропический Платицериум на ветвях дерева
Как размножаются папоротники в природе
Размножение папоротников стало предметом изучения не только ботаников и растениеводов, о споровом, вегетативном, бесполом способах размножения папоротников со схемами и описаниями рассказывают ученикам 5-6 классов в средней школе.
Споровое размножение Polypodióphyta подразделяется на два цикла. В них принимают участие два генетически единых вида одного и того же растения. На первом этапе из корневищ вырастают вайи, на нижней поверхности побегов формируются спорангии, в них образуются споры. Кучки спорангий со спорами собраны в мешочки (сорусы). Они хорошо видны на нижних сторонах вай. Споры из них высыпаются на землю после созревания.
Сорусы с созревшими спорангиями на обратной стороне вайи страусника
На втором этапе из спор на поверхности земли образуется небольшая, тонкая растительная пластина — заросток. В нем находятся разделенные между собой растительными волокнами сперматозоиды и яйцеклетки. Соединиться в единую клетку они могут лишь под внешним воздействием воды.
Соединиться в единую клетку они могут лишь под внешним воздействием воды.
Во время дождей созревшие мужские клетки подплывают к яйцеклеткам и образуют зиготы. Самая сильная из них формирует зародыш. Зародыш черпает силы из заростка, который постепенно отмирает. Зародыш формирует корневую систему, дает жизнь новому растению.
Обратите внимание! Корневище Polypodióphyta — это подземная часть стебля, в котором растения накапливают питательные вещества.
У некоторых видов папоротников споры образуются не на всех вайях. У папоротника Страусиное перо споры образуются в спорангиях, расположенных на специальных вайях, собранных в центре растения. Они отличаются размерами, толщиной тканей, собраны в пучок. Некоторые принимают эти ветви за цветоносы.
Страусник с центральными вайями
Как размножается папоротник в домашних условиях
Комнатное размножение и выращивание Polypodióphyta возможно несколькими путями — спорами, отсаживанием заростков и вегетативными способами:
- разделением корневищ;
- укоренением побегов;
- отделением и укоренением выводковых почек.

Самыми распространенными способами размножения являются споровый и прием разделения корневищ и корневых отростков. В условиях открытого грунта для размножения папоротниковидных культур в основном используют разделение кустов во время пересадки на новое место выращивания.
Строение корневой системы и поверхностных побегов Polypodióphyta
Как размножить папоротник разделением куста
Разделение корневых отростков и корневищ растений в саду проводят весной или ранней осенью. Комнатные разросшиеся папоротники можно размножать весь теплый период, совмещая пересадку с делением куста.
В условиях сада предварительно готовят участок для новых деленок. Для папоротников лучшим местом для выращивания будет участок сада, где растения смогут находиться в полутени.
Важно! Яркое солнце и густая тень не подходят папоротникообразным культурам.
Грунт для растений должен быть рыхлым, плодородным, хорошо пропускающим влагу. Специальную почвосмесь можно приобрести в садовом магазине или составить самостоятельно из листовой земли, песка и торфа (соотношение 2:1:1). Для комнатных Polypodióphyta на дно горшков насыпают дренажный слой. Почву перед пересадкой растений увлажняют.
Для комнатных Polypodióphyta на дно горшков насыпают дренажный слой. Почву перед пересадкой растений увлажняют.
Землю в горшке с растением, которое собираются рассаживать, увлажняют за 2 дня до пересадки. Начинают процедуру с того, что емкость с папоротником переворачивают набок и начинают катать по плоской поверхности, таким образом отделяют земляной ком от стенок горшка.
Удерживая одной рукой растение, горшок переворачивают вверх дном, вращательными движения вытаскивают куст вместе с комом земли. Осматривают состояние корней, розеток. Куст делят таким образом, чтобы в каждой деленке оставалось 2-3 розетки с хорошо развитыми корнями и почками роста.
Обратите внимание! Растениям для хорошей приживаемости необходимо обеспечить полутень, регулярные поливы, температуру воздуха в пределах 10-20 °С.
Разделение куста
Как размножить папоротник комнатный спорами
Многие цветоводы относятся к спорам Polypodióphyta, как к семенам обычных комнатных и садовых цветов. Споры у разных видов папоротников созревают в разные сроки. Признаком того, что споры можно использовать в качестве посадочного материала, становится изменение цвета спорангий на коричневый или бурый.
Споры у разных видов папоротников созревают в разные сроки. Признаком того, что споры можно использовать в качестве посадочного материала, становится изменение цвета спорангий на коричневый или бурый.
Спорангии вместе со спорами срезают с вайи и помещают в бумажный пакет. Приступать к посеву можно после того, как споры полностью высохнут.
Внимание! Они начнут высыпаться на дно пакета в виде коричневого порошка, поэтому с пакетом нужно обращаться осторожно, чтобы не рассыпать семенной материал.
Для посадки готовят неглубокие посадочные ящики и горшки с дренажными отверстиями или слоем керамзита, мелких кусков пенопласта. В ящики укладывают почвосмесь с добавлением крупнозернистого речного песка, листовой земли, торфа. Споры могут хорошо прорасти и в почве для фиалок.
Грунт хорошо увлажняют за несколько дней до посадочных работ. Перед посадкой землю утрамбовывают, споры располагают по поверхности грунта, ничем не присыпают. На посадочные емкости надевают прозрачные колпаки из полиэтиленовых пакетов или закрывают кусками стекла, то есть создают условия мини-тепличек. Периодически почву необходимо увлажнять, ящики проветривать.
Периодически почву необходимо увлажнять, ящики проветривать.
Важно! Почва в емкости с посеянными спорами должна быть постоянно увлажнена, иначе процесс оплодотворения яйцеклеток не произойдет.
Процесс прорастания спор является очень длительным. Первый этап развития спор — это появление заростков в виде мха. Корневая система начнется формироваться через несколько недель у образовавшегося зародыша, а до тех пор заростки получают влагу и питание через ризоиды — волоски, образовавшиеся на них после прорастания спор.
Ризоиды в нижней части заростков
Быстрый процесс размножения папоротников
Быстро размножать папоротники можно с помощью отделения выводковых почек и боковых побегов.
Боковые побеги — это тонкие усы, которые формирует папоротник наряду с вайями. Их назначение — обеспечить основное растение питательными веществами. Размножиться усами растение способно самостоятельно. При соприкосновении с влажным грунтом побеги врастают в него, пуская корни.
Выводковые почки образуются у некоторых видов Polypodióphyta в пазухах черешков листьев. В природных условиях они опадают, укореняются во влажной почве. В домашних условиях вайю с выводковыми почками пригибают к грунту, пришпиливают V-образным креплением, присыпают влажной почвой. Почки выпускают побеги. Когда на побегах сформируется 3 молодых листика их рассаживают в индивидуальные емкости.
Выводковые почки в пазухах черешков
Уход за проросшими папоротниками
Молодым растениям обеспечивают рассеянное освещение, оптимальную температуру воздуха, регулярные поливы отстоянной водой, подкормки.
Куст, находящийся на южной стороне дома, будет страдать от солнечных ожогов, а в тени — изменит окраску вай на бледно-зеленую. Оптимальным вариантом станет размещение растения таким образом, чтобы солнце попадало на него утром и вечером, а днем оно находилось в полутени.
В помещении, где находится Polypodióphyta, поддерживают влажность воздуха в пределах 70 %, при необходимости увлажняют. Температура воздуха не должна опускаться ниже 16 °С и подниматься выше 23 °С.
Тропические виды растений подкармливают специальным комплексным удобрением для папоротников два раза в год — весной и осенью, холодостойкие только весной. Поливы проводят после того, как подсохнет верхний слой грунта.
Важно! Температура воды должна быть комнатной.
Папоротники никогда не зацветут ни в квартире, ни на приусадебном участке, как бы хорошо за ними не ухаживали, поскольку просто не способны к этому. Но их замечательная густая шапка из ажурных листоподобных побегов украсит и садовый участок, и квартиру, и офис не хуже цветущего кустарника.
Размножение папоротников спорами, делением корня и вегетативно: советы и рекомендации
Главная — Ботаника — Комнатные растения — Как размножаются папоротники
Папоротники — самые древние растения, которые произрастают на Земле с древних времен. В мире насчитывается приблизительно 11 тысяч видов папоротников. В домашнем цветоводстве растения используют благодаря их красивым листьям. Они не цветут. В комнатных условиях выращивают тропические и субтропические растения, на улице для посадки используют кусты с умеренных широт.
В мире насчитывается приблизительно 11 тысяч видов папоротников. В домашнем цветоводстве растения используют благодаря их красивым листьям. Они не цветут. В комнатных условиях выращивают тропические и субтропические растения, на улице для посадки используют кусты с умеренных широт.
Богатая «шевелюра» растения компенсирует отсутствие цветов.
Строение и развитие папоротников
Папоротники состоят из корневой системы и листьев. Существует две разновидности папоротников:
- спорофиты — бесполые папоротники;
- гаметофиты — половые папоротники (мужские и женские).
Спорофиты представляют собой растения из субтропических и умеренных широт, у них листовая пластина изначально свернута в спираль. Именно эти растения чаще содержат в домашних условиях. Постепенно листы разворачиваются. Формы и размеры листов у каждого растения отличаются, они очень разнообразны. У некоторых видов длина листа достигает 6 метров.
Размах листьев у взрослого папоротника может достигать значительных размеров.
Растут листья у растения очень медленно. Развивается листовая почка под землей в течение двух лет. Только на третий год улиткообразная листовая пластина выводится наружу, а листовая почка к осени погибает. Вайя растет не от основания листа, а верхушкой. Именно вследствие длительного роста верхушкой длина вайи достигает крупных размеров.
Перед появлением подобной «улиточки» должно пройти немало времени.
Листы необходимы растению для фотосинтеза. На нижней стороне пластины у них формируются споры. У некоторых видов споры образуются на специальных спороносных листьях, у других — на листах в средней части вайи или ее верхушечной части. У разных видов существуют различные способы размножения. Папоротники могут размножаться делением корня или выводковыми почками, которые формируются на листьях. Некоторые разновидности папоротников ядовиты.
Если вы хотите создать на дачном участке райский уголок, без флоксов вам не обойтись. Эти яркие, пышные цветы смотрятся очень нарядно, с их помощью можно создавать настоящие композиции.
Драцена — растение, приносящее в дом удачу и счастье. Ее очень просто выращивать в домашних условиях, растение не прихотливо, не требует особого ухода. Как размножается драцена рассказано тут.
Виды домашних папоротников
Для комнатного разведения в горшках подходят такие виды папоротников:
- Адиантум (Adiantum) — изящное растение с перистыми вайями, расположенными на тонких темно-коричневых черешках. Пышная крона формируется из мелко рассеченных листьев. Он быстро растет, весьма неприхотлив, хорошо переносит затемненные места.
Данный вид папоротника легко переносит нехватку солнечного света.
- Асплениум (Asplenium) привлекает эффектными, блестящими листьями светло-зеленого цвета. Растение отличается тем, что листья не разделяются на сегменты и формируют величественную форму. По центру листа проходит центральная жилка, которая через время темнеет и становится выразительнее.
Глянцевые, цельные листы папоротника выглядят очень эффектно.
- Нефролепис (Nephrolepis) — неприхотливое растение.
 Различают несколько видов нефролеписов, которые отличаются формой и размером вай. Ажурные листья светло-зеленого цвета собраны в розетку. Длина вай достигает 1 м. На нижней стороне вай расположены сорусы со спорами, которые необходимы растению для размножения.
Различают несколько видов нефролеписов, которые отличаются формой и размером вай. Ажурные листья светло-зеленого цвета собраны в розетку. Длина вай достигает 1 м. На нижней стороне вай расположены сорусы со спорами, которые необходимы растению для размножения.Нефролепис может похвастаться тонкими, воздушными листьями.
- Платицеритум (Platycerium) отличается тем, что создает вайи двух типов. Листы заметно различаются по форме и назначению: спороносные вайи и стерильные вайи округлой формы. Длина вай достигает 50—70 см, листы нельзя протирать, иначе нарушится серебристая опушка.
Размножение папоротников спорами
Цикл размножения папоротников включает сбор созревших спор, хранение их для дозревания и высадку в грунт. Споры у растений размещены в спорангиях, расположенных точечно — в сорусах. При созревании спорангии становятся коричневыми. Каждая разновидность имеет свою скорость роста и особенности прорастания, поэтому лучше разграничивать посевы спор разных сортов. У созревших спор сорусы буреют. Споры срезают с участками вайи и хранят в бумажном пакетике для дозревания и просушивания. Пакетик нужно тщательно запечатать, так как споры легко летают. Открывают его только перед закладкой в почву.
У созревших спор сорусы буреют. Споры срезают с участками вайи и хранят в бумажном пакетике для дозревания и просушивания. Пакетик нужно тщательно запечатать, так как споры легко летают. Открывают его только перед закладкой в почву.
Коричневые мешочки на листьях и есть будущие молодые папоротники.
Чтобы споры отделились от листа, необходимо просто постучать по пакету или хорошо его потрясти. Созревшие споры имеют вид коричневого порошка. Для посева применяют такие виды грунтовой смеси:
- песок и верховой (бурый) торф в соотношении 1:3;
- песок, листовую землю верховой торф и в соотношении 1:1:2;
- песок и листовую землю в соотношении 1:3;
- готовую земляную смесь для фиалок.
Субстрат старательно просеивают, чтобы убрать крупные и твердые частицы. До посева почву рекомендуется простерилизовать посредством пропаривания для уничтожения сорняков и вредителей в течение 4 часов. В качестве горшка используют неглубокую емкость из пластика или стекла. Контейнер наполняют влажным подготовленным грунтом и слегка его утрамбовывают. Толщина слоя составляет не более 4 см.
Контейнер наполняют влажным подготовленным грунтом и слегка его утрамбовывают. Толщина слоя составляет не более 4 см.
Сбереженные в пакетике споры засеивайте в неглубокий контейнер.
Споры плотно и равномерно распределяют по поверхности и не присыпают. Это бесполое размножение. Поверхность прикрывают стеклом, чтобы поддерживать высокую влажность. Контейнер ставят на свету, но защищают от солнца. Температура в комнате должна быть около +15—25°С. При низких температурах они не прорастают.
Период прорастания спор зависит от вида растения. Споры прорастают в интервале от нескольких недель до двух лет.
Весь период прорастания следует поддерживать стабильную влажность грунта, увлажняя сеянцы только с поддона.Вода для полива должна быть без хлора. Периодически поверхность грунта нужно проветривать, открывая стекло. Проросшие споры покрывают поверхность в виде зеленого мха. У пророщенных растений отсутствуют корни, и к почве они крепятся специальными волосинками — ризоидами.
Особенности ухода за проросшими папоротниками
Только через 1,5—2 месяца на ростках начинают прорастать корни и листья. Покровное стекло в этот период сначала убирают на 1—2 часа в сутки, увеличивая постепенно промежуток. Когда посевы раскрывают на постоянное время, их обязательно систематически опрыскивают водой. Рекомендуется обрабатывать ростки раствором эпина или микроэлементов каждые 2 недели 2—3 раза.
Регулярный полив и опрыскивания будут полезны для всходов.
Контейнер с ростками рекомендуется поместить в аквариум, который нужно держать закрытым. Постепенно растения адаптируют к открытому воздуху. У папоротников можно пикировать проросшие споры еще до формирования корня. Их размещают на поверхности грунта и тщательно опрыскивают водой. Растения со сформированными корнями пикируют, как обычно. Через 7—8 месяцев получаются хорошо развитые кусты.
Лучшая живая изгородь получается из барбариса. Этот кустарник необычно смотрится на участках рядом с хвойными деревьями или цветами, барбарис может размножаться семенами и черенками.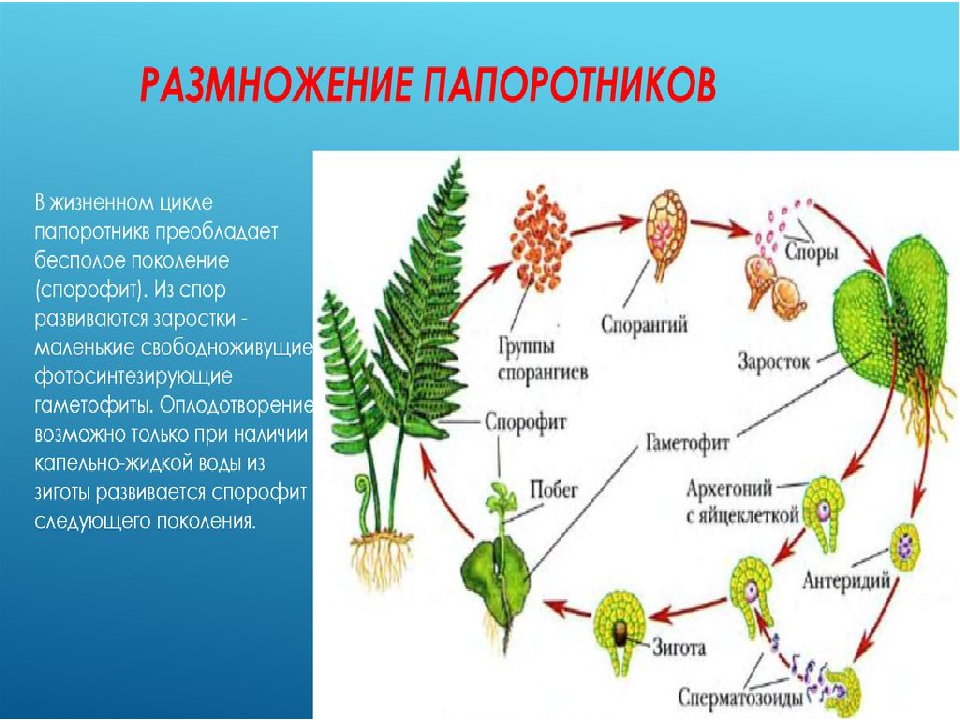 Ягоды барбариса содержат множество витаминов, каротин и успокаивающие вещества. Они незаменимы в процессе приготовления плова, супов и салатов.
Ягоды барбариса содержат множество витаминов, каротин и успокаивающие вещества. Они незаменимы в процессе приготовления плова, супов и салатов.
Волнистые попугайчики — веселые и забавные обители квартиры. Эти птицы могут стать настоящими друзьями. В этой статье мы поговорим о размножении волнистых попугайчиков.
Улитки являются своеобразными санитарами в аквариуме, они эффективно чистят стенки домашнего бассейна. Перейдите по этой ссылке www.razmnojenie.ru/zoologiya/mollyuski/ulitki.html и вы узнаете все о размножении улиток в домашнем аквариуме.
Вегетативное размножение папоротниковых
Вегетативным способом чаще всего размножаются садовые виды папоротников, некоторые комнатные растения и половые культуры. Среди комнатных видов наиболее пригодны для деления корня адиантумы, пеллеи, листовики, нефролеписы, птерисы. Растения размножают осенью в процессе пересадки или весной, когда ваи еще не выросли.
Вегетативный способ размножения пригоден не для всех видов папоротниковых.
Для деления не подходят растения, которые имеют только одну точку роста. Корневую систему ножом аккуратно разрезают между розетками. На каждом разделенном растении должна остаться часть корневой системы. Процедуру деления выполняют только в прохладную погоду. Растения высаживают в грунт и тщательно поливают. При размножении осенью растения также систематически поливают.
Необходимо учесть температурные условия при делении.
Некоторые виды размножаются вегетативно, посредством выводковых почек на черешках листьев. Выводковые почки пересаживают в тепличку, например, в аквариум. В тепличке поддерживают постоянную влажность. Когда растения укоренятся, их с частью грунта пересаживают на постоянное место. Растений, которые размножаются таким способом, очень мало, одно из них — пузырник луковиценосный.
Размножение папоротников
☰
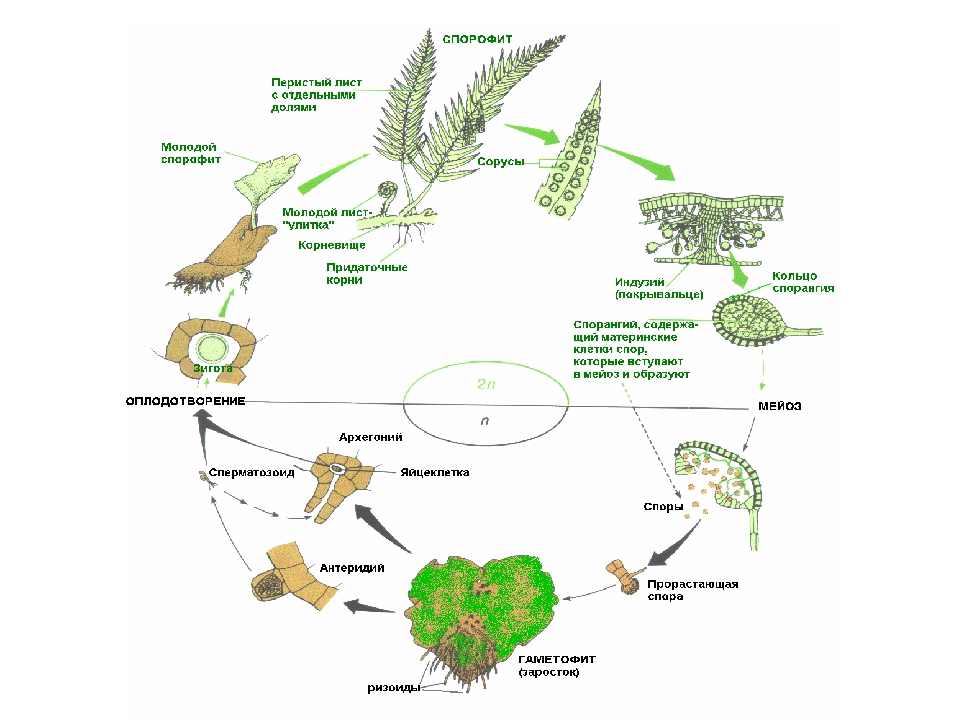
У папоротников, как и у мхов, в жизненном цикле чередуются два поколения: гаметофит и спорофит. Однако, в отличие от мхов, у папоротников преобладает спорофит. То есть растение, которое мы в обиходе и называем папоротником, представляет собой спорофит. В клетках спорофита двойной набор хромосом, а в клетках гаметофита — одинарный.
Однако, в отличие от мхов, у папоротников преобладает спорофит. То есть растение, которое мы в обиходе и называем папоротником, представляет собой спорофит. В клетках спорофита двойной набор хромосом, а в клетках гаметофита — одинарный.
Можно считать, что основное размножение папоротников происходит с помощью спор, так как именно благодаря им происходит увеличение количества новых растений папоротника, а также расселение. При половом размножении (с помощью половых клеток) новое растение вырастает там, где уже был заросток, выросший из споры.
На вайях (листьях папоротника) с нижней стороны образуются бугорки, состоящие из спорангиев. В них развиваются споры, содержащие одинарный набор хромосом (т. е. они уже являются гаметофитом, развивающемся на спорофите). Созревшие споры высыпают и разносятся ветром и водой.
Попав в благоприятные условия, спора прорастает в так называемый заросток, который является гаметофитом. Заросток папоротника выглядит как маленькая зеленая пластинка (в несколько миллиметров), но она способна к фотосинтезу, а значит, питается самостоятельно. У заростка есть корнеподобные образования (ризоиды), которыми он прикрепляется к почве и всасывает воду с минеральными веществами.
У заростка есть корнеподобные образования (ризоиды), которыми он прикрепляется к почве и всасывает воду с минеральными веществами.
На нижней стороне заростков образуются половые клетки: сперматозоиды и яйцеклетки. Они гаплоидны, так же как заросток. Когда под заростками скапливается вода, сперматозоид может подплыть к яйцеклетке и оплодотворить ее.
В результате оплодотворения образуется зигота. Поскольку в ней объединяются хромосомные наборы сперматозоида и яйцеклетки, то она имеет двойной набор хромосом, т. е. представляет собой спорофит.
Зигота постепенно развивается в зародыш. При этом зародыш на начальных стадиях своего развития остается связанным с заростком и питается за его счет. Когда у зародыша появляется собственный корень и зеленые части, он начинает питаться самостоятельно.
Зародыш вырастает во взрослое растение папоротника (спорофит).
Папоротники способны также к вегетативному размножению. Оно может осуществляться с помощью корневищ, выводковых почек и др.
Оно может осуществляться с помощью корневищ, выводковых почек и др.
схема, особенности полового и вегетативного размножения
Папоротники относятся к древнейшей группе растений, существовавшей задолго до появления цветущей растительности и сохранившихся до сих пор. Сегодня папоротник – один из популярных растений для оформления ландшафтного дизайна. В этой теме рассмотрим размножение папоротников в домашних условиях, а также их разновидности и развитие.
Содержание материала:
Виды и сорта папоротников
Существует огромное количество групп и сортов папоротника.
Асплениум.
Известен любовью к скалистым поверхностям. Формирует отдельное семейство. Описание следующее: округлые, кожистые листовые пластинки на тонких веточках. Самые известные два вида: постенный и северный. Первый часто встречается на стенах старых каменных построек. Растет преимущественно в каменных расщелинах. Северный любит скалистые места северной Европы и Азии.
Вудсия.
Относится к семейству асплениевых. Низкорослый папоротник, отличающийся «пушистой» листвой – из небольшого корня вырастают тонкие листья, от которых в две стороны расходятся листочки меньшего размера. Имеет несколько сортов (эльбская, многорядниковая), которые отличаются размером и плотностью листочков.
Кочедыжник.
Также относится к семейству Асплениума. Эта разновидность имеет около 200 сортов. Из них наиболее известны женский, китайский-красноватый (с красными прожилками). Описание: высокий кустарник с ажурными, легкими листовыми перьями.
Многорядник.
Как и предыдущие группы, входит в семейство Асплениума. Отличается от предыдущих шириной листовых веток, любовью к густой тени лесистой местности. Известны такие сорта: многорядник Брауна, трехраздельный, щетинковый.
Это интересно: папоротник комнатный
Орляк.
Относится к семейству циатейных. Популярен своей неприхотливостью – сорта этой группы встречаются на всех континентах нашей планеты: тундра, пустошь, лесистые заросли. Его нет разве что в пустыне и степях. От других разновидностей отличается огромным размером листовых ветвей – до полутора метров.
Его нет разве что в пустыне и степях. От других разновидностей отличается огромным размером листовых ветвей – до полутора метров.
Осмунда.
Формирует собственное семейство. Один из самых древних видов папоротниковых. В далекие времена произрастали на всех материках, но сегодня встречаются лишь на Кавказе, в лесах Восточной Азии и Северной Америки. Сорта известны следующие: азиатская, Клейтона, королевская. Любит небольшую тень и болотистую местность.
Скребница.
Относится к асплениумному семейству. Ее отличительная особенность – уникальная в своем семействе любовь к суше и большому количеству света. Листья имеет кожистые, плотные, в отличие от сородичей. Предпочитает известняковые скалы.
К распространенным разновидностям папоротника относятся Страусник (форма листьев напоминает страусиные перья), Телиптерис (отличается буйной пушистостью зелени), Фегоптерис, Щитовник, Онколея.
Факт. Именно папоротники много тысяч лет назад имели в своем роде сорта древесных растений.
Деревья со временем отмирали, а их древесина уходила вглубь почвы, спрессовывалась и дожила до наших дней в виде каменного угля.
Строение и развитие папоротников
Главная фаза жизненного цикла – спорофит.
Особенности строения спорофита:
- не длинные корни, отходящие от основного продолговатого корневища. Первичный корень быстро отмирает, а небольшие корешки крепнут, на них образуются почки;
- побеги – образуются из почек на корнях, являются главным способом размножения папоротника;
- листья – вырастают из корня;
- спорангии, собранные в небольшие пучки с названием «сорусы» – маленькие зеленые пластинчатые коробочки со спорами – второй способ размножения папоротника.
Жизненные циклы:
- Бесполое растение. Полноценный папоротниковый куст, производящий семена.
- Споры. Семена из коробочки, попавшие в почву.
- Заросток. Небольшое зеленое растение, прикрепленное к почве, производящее сразу мужские и женские клетки.

- Гаметы. Стадия размножения, слияния разнополых клеток.
- Зародыш. Формирование нового бесполого растения.
На заметку. Зеленую массу папоротника не принято называть листьями, так как по строению она совершенно не схожа с привычной листвой цветущих кустарниковых растений.
Схема и описание размножения растений
Размножение папоротника происходит при помощи семян.
Схема размножения папоротника происходит таким образом: с нижней стороны листа папоротника находятся сорусы, которые при созревании отделяются от листовой пластины. Спорангии в кольце соруса разрываются, рассеивая вокруг себя споры папоротника. Затем из споры образуется небольшая зеленая пластинка, прорастающая в почву. Этот заросток крепится к почве ризодами, имеет растительные половые органы мужские и женские. Как только на пластинку попадет вода, например, после дождя, происходит оплодотворение: в женских органах (архегониях) есть яйцеклетка, к которой по каплям воды передвигаются из мужских органов (антеридий) сперматозоиды, и оплодотворяют ее. Получается полноценный зародыш, прикрепленный к заростку, от которого питается во время развития. Далее со временем вырастает растение.
Половое размножение и его особенности
Для собственноручного размножения папоротника в домашних условиях необходимо собирать споры, освободившиеся от оболочки и высеивать в нужном месте. Этот участок должен быть хорошо увлажненным. Условие для благополучного зачатия нового растения – высокий уровень влажности.
Для выращивания рекомендуется использовать смесь из торфа, простерилизованной почвы и древесного угля в соотношении 8:2:1 соответственно. Полученной смесью заполняют небольшой горшок почти до верха, затем ее хорошенько утрамбовывают, посыпают кирпичной крошкой. В эту крошку разбрасывают камни. Сверху горшок плотно накрывают прозрачным стеклом. Всю конструкцию ставят в поддон с отстоянной водой. Температура выращивания 21 градус. Место должно быть затемненным.
Первые заростки появляются через месяц, еще через два – оплодотворенные маленькие папоротники пускают первые листочки.
Вегетативное размножение растения
Вегетативное размножение – один из способов размножения папоротника путем деления, применяющийся садоводами. В природе такой способ не может происходить, да и подходит он не всем видам папоротниковых.
Важно! Для деления куста необходимо соблюдать условие – на одном корневище должно быть несколько розеток. Тогда их разделяют на несколько кустов.
Сроки деления – август и сентябрь, когда погода уж стала прохладной, либо весной, до раскрытия листовой вайи. Растение нужно выкопать и аккуратно ножом разрезать корневище между розетками, после чего посадить на новые места и некоторое время обильно поливать.
Цикл размножения
Цикл размножения делится на две эпохи:
- Бесполое поколение. Точкой отсчета является зигота, из которой произрастает молодой спорофит. Далее он развивается в полноценное взрослое растение, на листьях которого формируются спорангии.
- Половое поколение. Наступает в момент раскрытия спорангии и попадания спор в грунт. Зрелые споры прорастают небольшими «ножками» в почву, формируется в зрелого гаметофита. На этом этапе женские и мужские клетки готовы к слиянию. Далее при удобных условиях они сливаются, и цикл снова переходит на этап бесполого поколения.
Папоротник довольно распространенное растение при оформлении садового участка. Его не обязательно покупать в магазине – можно выкопать в лесистых зарослях. Главное: кусты нужно брать на участке, где папоротниковые хорошо размножились, то есть образовали обширные заросли. На черешках должна быть хотя бы одна розетка, почка, более-менее развитая корневая система, чтобы растение могло адаптироваться на новом месте. Место посадки должно быть влажным, без доступа прямых лучей солнца.
Размножение папоротников семенами, спорами, корневищем, способы размножения
Размножение папоротников – процесс разведения спорового декоративного растения в домашних условиях. Изначально его считали диким растением, которое произрастает исключительно в естественных условиях. Сегодня множество дачников занимаются разведением папоротников, чтобы создать привлекательный ландшафтный дизайн садового участка. Из 11 тысяч видов одомашненными считают только 2000 разновидностей, которые размножают не только естественным образом, но и рассадой, отростками.
Как происходит размножение папоротников в природе
Обычно в естественных условиях папоротники размножаются с помощью спор или выводковой почки. В течение всего жизненного цикла растения проходят стадии спорофит и гаметофит. Иногда при распространении корневой системы и увеличении новых организмов происходит самостоятельное расселение через половые ветки. В подобных местах появляется заросток, который зарождается на месте спорового кармана.
Процесс размножения прост: на вайях образуются спорангии, в которых развиваются споры с одинарным набором хромосом. После созревания споры распространяются по воде или ветру. Папоротник размножается спорами только в благоприятных температурных условиях. Попадая в подобные условия, заросток прорастает, затем закрепляется на поверхности с помощью ризоидов. В последующем в нижней части пластины развиваются половые клетки со сперматозоидами и яйцеклетками. После полного созревания происходит оплодотворение и зарождение зиготы. Зародыш питается от заростка, пока не обретет собственные корни. Таким образом, из зародыша или гаметофита вырастает настоящий кустарник.
Как можно размножить папоротник в саду
В садоводстве используют несколько видов размножения папоротника: деление куста, рассада, споры, корневые почки. Ранее делали сбор спорангий с целью дозревания спор в домашних условиях. Процесс разведения заключался в подготовке посадочного материала, который хранят в плотно запечатанных конвертах и сажают во влажный грунт.
После посадки лоток с семенами накрывают плотным полиэтиленом или стеклом, чтобы создать необходимый микроклимат. Первые всходы появляются через 20-30 дней. Поросль из ризоидной зелени обрабатывают 3 раза в день раствором эпина. Проталлиум или папоротниковую рассаду высаживают в отдельные торфяные стаканы, которые будут поглощать излишки влаги при поливе. Когда ростки достигают 5-10 мм, рассаду пикируют. За 6 месяцев проводят 3 пересадки, затем в возрасте 8 месяцев папоротник высаживают в клумбу или цветочный горшок.
Уличное растение рассаживают путем деления куста, который откапывают в начале весны. Заранее копают лунки, затем делят главный куст на несколько небольших частей. Перед посадкой обрезают гнилые части растения и присыпают землей. Дочерние кусты начинают развиваться после недели акклиматизации на новой земле. Эта особенность размножения папоротников заметна по недельной вялости ветвей, которая уходит после укоренения.
Внимание! Рассадный способ посадки и деление основного куста для размножения папоротника считаются самыми эффективными.
Посадочный материал в виде спор не всегда можно собрать вовремя или приобрести качественный товар в магазине. Однако и рассада может не пойти в рост, если не обеспечивать комфортные условия микроклимата и грунта.
Какие условия необходимы для размножения папоротников
В основном комфортные условия для благоприятного развития растения – высокая контролируемая влажность в помещении или увлажненный грунт на улице. Оптимальное время для начала вегетативного размножения папоротника – ранняя весна с постоянной плюсовой температурой воздуха. Рассадку кустов также делают летом после дождя, когда земля не требует дополнительного увлажнения. Папоротник не доминирует над сторонними растениями, поэтому может соседствовать с различными видами кустарников.
Чем реже садовод планирует поливать растение, тем дальше в тень сажают кусты. Растение укореняется и хорошо развивается при любом виде освещения. При посадке на солнечной стороне участка необходимо следить за состоянием куста и грунта. Быстро вянущие ветви – явный показатель нехватки влаги и витаминов. Эффективный полив вроде летнего душа после заката из мелкой лейки сделает папоротник пышным и ярким. Стоит заметить, что в тени папоротники более ветвистые, тогда как на солнце растут компактными кустами со светлой зеленью.
Как размножить папоротник делением куста
Результативный способ размножения кустарника, который можно делать в любой теплый сезон года, заключается в делении куста. Для начала за день до рассадки корни растения обильно заливают водой. По описанию и схеме размножения папоротники сажают на глубину до 20-30 см, хотя посадочную яму копают вглубь на 50-70 см. Дно засыпаю щебнем, смешанным с субстратом и удобрениями. Кусты делят на 4 небольшие части так, чтобы корни сильно не травмировались. На каждой части растения должно остаться 2 или 3 розетки. Корневища без почек роста будут долго приживаться или могут не укорениться.
По субстрату на дне лунки аккуратно расправляют корни, затем присыпают землей. После посадки папоротник поливают и опрыскивают разведенным раствором фитонцидов от насекомых. В первый год роста делают прикорневой круг для полива и мульчируют корни сеном или крупными опилками. Если листья начинают желтеть или покрываться ржавчиной, нужно произвести подкормку компостом или минеральным удобрением. Способы полива нужно чередовать: душ из лейки 2 раза в неделю и 1 прикорневой полив. Пикировку делают только по необходимости, если куст не принимается или почва слишком тяжелая, кислая.
Важно! Папоротник с наличием 1 корневой розетки делить нельзя.
Как размножить папоротник спорами
Выращивание папоротников из спор – довольно трудоемкий процесс возделывания кустарника, за которым нужен особый уход до момента первой пересадки. Посадочный материал приобретают в фирменных магазинах, хотя процессом сбора спорангий можно заняться самостоятельно.
Споры можно высаживать в любое время года, если посадка производится для выращивания комнатного папоротника. Уличные споровые растения сажают ранней осенью или весной. Споры рассыпают по поверхности влажного грунта, затем присыпают слоем земли в 3-4 см. Почку опрыскивают из пульверизатора и накрывают стеклом, пищевой пленкой так, чтобы внутри собирался конденсат. После появления первых всходов покрытие снимают в дневное время, а когда появляется проталлиум, делают рассадку по горшкам.
Важно! Споры сажают исключительно в стеклянные емкости, поливают отстоявшейся водой без хлора.
До образования первых листьев рассаду держат под стеклом и открывают на 2-3 часа. При частом и умеренном поливе 2-3 раз в неделю ростки быстро пойдут в рост. В помещении необходима постоянная плюсовая температура + 20-23 °С. Дикие сорта более приспособлены к экстренным условиям, но для сбора спорангий в лесу сложно подгадать время. Это не самый эффективный способ размножения папоротника, но при обеспечении должного ухода и подготовки посадочного материала можно вырастить здоровое растение.
Можно ли вырастить папоротник из семян
Ни в коем случае нельзя путать споры папоротника с семенами. Посадочный материал готовят самостоятельно. Как только на нижних листах образуются спорангии, делают обрезку нескольких веток. Споровые мешки не успеют раскрыться, а когда дозреют, то будут готовы к сушке. Семена снимают с листов и сушат под марлей в помещении с низкой влажностью. Размножать папоротник семенами начинают в середине марта или в конце апреля.
Семена высаживают в грунт, когда на ощупь их можно раскрошить. Способ выращивания кустарника почти ничем не отличается от спорового, за исключением того, что споры не всегда прорастают и большая часть посадочного материала погибает на стадии развития до проталлиума. В первые 2-3 месяца полив осуществляют 1-2 раза в неделю. Минимальная температура для посадки на улице допускается до + 10 °С, в комнате до + 15-18 °С. В возрасте 6 месяцев делают пересадку в новый грунт, подкармливают фосфатами. При возрасте 1-2 лет кусты делят на рассаду.
Заключение
Размножение папоротников – достаточно увлекательное и познавательное дело для любителей украсить собственный сад пышной зеленью. Растение неприхотливо к условиям произрастания, но требует пристального внимания на стадии разведения и выращивания после посадки. Своевременные подкормки и водные процедуры будут способствовать благоприятному развитию папоротника. Пышные и здоровые кустарники радуют взор дачников и любителей садоводства.
Строение и размножение папоротников — СПОРОВЫЕ РАСТЕНИЯ
Цели урока:
обучающая: изучить особенности строения, размножения и развития папоротников на примере щитовника мужского;
развивающая: развивать у учащихся умения устанавливать причинно-следственные связи;
воспитательная: формировать ценностное отношение к окружающей природе; на примерах показать уязвимость папоротников и необходимость осторожного обращения с растениями в лесу.
Тип урока: изучение нового материала.
Методы обучения: беседа, описание, “биологический” диктант, рассказ с демонстрацией гербарных экземпляров.
Формы деятельности учащихся: индивидуальная, фронтальная.
Опорные термины, понятия: жизненный цикл, спорофит, гаметофит, спора, архегонии, антеридии, оплодотворение, зигота.
Вводимые понятия: папоротники, вайи, сорусы, заросток.
Материалы и оборудование: гербарный материал, лупы, раздаточный материал, различные виды комнатных папоротников, таблица “Папоротник щитовник мужской”, компьютер, мультимедийный проектор, презентация, карточки с заданиями.
Перечень знаний, умений, навыков, которые предполагается развивать на данном уроке:
— характеризовать строение, размножение и развитие папоротников.
- Проверка домашнего задания
Ø Деятельность учителя
Проводит графический диктант, в котором необходимо отметить верные суждения знаком “+”, неверные — знаком “-”.
- Среди высших растений моховидные представляют собой обособленную и тупиковую ветвь развития. Они произошли около 350 млн лет назад от первых наземных растений псилофитов — потомков прибрежных водорослей.
- Мхи прикрепляются к почве с помощью корней, через которые они всасывают питательные вещества.
- При разработке торфа были найдены хорошо сохранившиеся останки рыцаря в доспехах. Это объясняется тем, что сфагновые мхи выделяют обладающие бактерицидными свойствами гуминовые кислоты, которые убивают бактерии, и гниение не происходит.
- Споры хвощевидных используют в медицине в качестве детской присыпки, в ветеринарии, а также в промышленности для получения желтой и зеленой красок.
- Важное условие полового размножения мхов, плаунов, хвощей — наличие воды.
Ø Деятельность учащихся
Предполагаемые правильные ответы: 1 — “+”; 2 — “-”; 3 — “+”; 4 — “-”; 5 — “+”.
Ø Деятельность учителя
Нескольким учащимся может предложить выполнить индивидуальные задания по карточкам:
Карточка № 1. Сделать подписи к рисунку “Строение сфагнума п кукушкина льна”:
Карточка № 2. В схеме “Размножение мха” вставить недостающие элементы:
Ø Деятельность учащихся
Выполняют индивидуальные задания, на которые каждому отведено по 5 минут.
Ø Деятельность учителя
Задает вопросы:
- Почему мхи называют “земноводными” в мире растений?
- Чем мхи прикрепляются к почве?
- Какое поколение доминирует в жизненном цикле мхов?
- Где находится половое и бесполое поколение у мхов?
- Из чего вырастает растение мха?
- Почему процесс образования торфа связан со сфагновым мхом?
Ø Деятельность учащихся
Предполагаемые правильные ответы:
- Клетки мхов легко теряют или поглощают воду. Для полового размножения им необходима водная среда.
- Ризоидами.
- Гаметофит.
- На одном растении.
- Из споры.
- Он задерживает влагу и не гниет в толще отложений.
- II. Организационный момент, постановка проблемы
Ø Деятельность учителя
Сообщает учащимся, что на уроке они продолжат изучение высших споровых растений, среди которых выделяются представители отдела Папоротниковидные. Это древнейшая группа настоящих наземных растений, известная с девонского периода. Сейчас известно около 12 тыс. папоротников. Демонстрирует слайд № 1.
Ø Деятельность учащихся
Слушают объяснение учителя.
Ø Деятельность учителя
Обращает внимание учащихся на тему урока и предлагает вспомнить и рассказать то, что им известно о папоротниках.
Ø Деятельность учащихся
Записывают в рабочих тетрадях тему урока.
Вспоминают все, что знают о папоротниках.
Ø Деятельность учителя
Знакомит учащихся с преданием: “Только один раз в году — в ночь на Ивана Купалу — в густом лесу в полночь раскрывается удивительный цветок папоротника”. Читает отрывок из произведения II. В. Гоголя “Ночь накануне Ивана Купалы”: “Глядь, краснеет маленькая цветочная почка и, как будто живая, движется. В самом деле, чудно! Движется и становится все больше, больше и краснеет, как горячий уголь. Вспыхнула звездочка, что-то тихо затрещало, и цветок развернулся, словно пламя, осветив и других около себя. Только смелый и удачливый человек мог заполучить цветок папоротника — разрыв-травы, от которого распадались все замки и запоры и сами в руки давались клады”.
Ø Деятельность учащихся
Слушают предание.
III. Изучение нового материала
Ø Деятельность учителя
Задает вопрос: “Можно ли верить преданию о цветке папоротника и его чудодейственной силе?”.
Ø Деятельность учащихся
Отвечают на поставленный вопрос. Выдвигают собственные гипотезы.
Ø Деятельность учителя
Рассказывает: “Чтобы ответить на поставленный вопрос, нам необходимо познакомиться со строением и жизнедеятельностью папоротников. Возникновение легенды связано с тем, что у некоторых видов папоротников, например у ужовника обыкновенного, одна часть листа выполняет функцию фотосинтеза, а другая — спороношения. Спороносная часть листа похожа на нераскрывшийся бутон цветка”. Демонстрирует слайд № 2.
Ø Деятельность учащихся
Записывают в тетрадях новые понятия.
Ø Деятельность учителя
Рассказывает, что тело папоротника делится на корень и побег, как у настоящих наземных растений, приспособленных к жизни на суше. Побег образован стеблем и листьями. Задает вопрос: “Как вы думаете, какую функцию выполняют корень и побег?”. Демонстрирует слайд № 3.
Ø Деятельность учащихся
Предполагаемый правильный ответ: Корень обеспечивает растение водой и минеральными солями, а побег синтезирует органические вещества.
Ø Деятельность учителя
Сообщает, что листья папоротников очень специфичны и называются “вайи”. Для них характерен длительный рост вершины, проявляющийся в образовании улитки. Задает вопрос учащимся: “Какую функцию, по вашему мнению, выполняют листья и стебель папоротников?”.
Ø Деятельность учащихся
Записывают определение понятия “вайи” в рабочих тетрадях.
Предполагаемый правильный ответ: Основная функция листьев — фотосинтез и спорообразование, а стебля — транспорт веществ и вынос листьев на поверхность.
Ø Деятельность учителя
Задает вопрос: “Как вы думаете, какие особенности строения листьев связаны с выполнением названных функций?”.
Ø Деятельность учащихся
Предполагаемый правильный ответ: С фотосинтезом связано наличие хлоропластов, обусловливающих зеленую окраску листьев, и широкой листовой пластинки, обеспечивающей более полное использование света под пологом леса. Спороношение обеспечено наличием спорангиев на нижней поверхности листьев.
Ø Деятельность учителя
Обращает внимание учащихся на то, что зародышевый корень у папоротников быстро отмирает и на смену ему появляются придаточные корни, возникающие на стебле.
Ø Деятельность учащихся
Записывают в рабочих тетрадях следующий факт: “Корень, как орган минерального питания, поглощает из почвы воду и минеральные соли, которые по проводящей системе корня, стебля и листа поступают во все органы папоротника. Сам корень использует готовые органические вещества, синтезируемые в листьях и поступающие ко всем органам растения по проводящим тканям — ксилеме и флоэме”.
Ø Деятельность учителя
Ставит перед учащимися вопрос: “Каким образом происходит размножение и развитие папоротников?”. Затем сообщает, что развитие папоротников происходит со сменой поколений спорофита и гаметофита. Демонстрирует слайд № 4.
Ø Деятельность учащихся
Отмечают в рабочих тетрадях особенности размножения папоротников.
Ø Деятельность учителя
Осуществляет актуализацию опорных знаний учащихся. Предлагает ответить на вопросы:
- Какое поколение называют гаметофитом?
- Какое поколение называют спорофитом?
- Чем представлено половое поколение мхов?
- Что собой представляет бесполое поколение мхов?
- Какое поколение преобладает в жизненном цикле мхов?
Ø Деятельность учащихся
Предполагаемые правильные ответы:
- Гаметофит — половое гаплоидное поколение, на котором формируются половые клетки: яйцеклетки и сперматозоиды.
- Спорофит — бесполое диплоидное поколение, на котором формируются споры.
- Половое поколение мхов представлено зеленым травянистым растением.
- Бесполое поколение мхов представляет собой коробочку на ножке, живущую на спорофите.
- В жизненном цикле мхов преобладает гаметофит.
Ø Деятельность учителя
Следующим вопросом, который необходимо рассмотреть на уроке, является изучение жизненного цикла папоротников.
Отмечает, что чередуются бесполое и половое поколения — спорофит и гаметофит. Преобладает фаза спорофита — именно его мы называем папоротником. На нижней стороне листа можно заметить маленькие округлые образования. Демонстрирует слайд № 5.
Объясняет учащимся: “Отдельный спорангий трудно увидеть простым глазом. Его строение идеально приспособлено для разбрасывания спор. Зрелые споры высыпаются и разносятся ветром. Из споры вырастает крошечное растеньице, которое похоже на сердцевидную пластинку. Это — заросток, являющийся гаметофитом. Заростки при внимательном осмотре можно обнаружить на поверхности лесной подстилки. На нижней стороне заростка имеются ризоиды. Там же образуются антеридии и архегонии, в которых развиваются половые клетки — соответственно подвижные сперматозоиды и неподвижные яйцеклетки. Сперматозоиду, чтобы доплыть до яйцеклетки, необходима вода. После слияния половых клеток образуется одна клетка — зигота. Из зиготы развивается зародыш, который вырастает во взрослое растение — спорофит (сам папоротник)”.
Ø Деятельность учащихся
Составляют опорную схему жизненного цикла папоротников и рисуют ее в рабочих тетрадях.
- Закрепление изученного материала
Ø Деятельность учителя
Предлагает учащимся проанализировать записи, которые были сделаны в рабочих тетрадях в течение урока, и сформулировать вывод: что нового они узнали о папоротниках?
Ø Деятельность учащихся
Формулируют устно ответ на поставленный вопрос.
Ø Деятельность учителя
Предлагает выполнить дифференцированные задания:
- Сделайте подписи к рисунку “Строение папоротника”.
- В предложенных карточках учащимся необходимо подчеркнуть правильные ответы.
Пример карточки:
- Что развивается из спор папоротника? (Гаметофит; спорофит.)
- Чем представлен гаметофит папоротника? (Травянистым растением с корневищем, стеблем и листьями; коробочкой со спорами; заростком.)
- Чем представлен спорофит папоротников? (Травянистым растением с корневищем, стеблем и листьями; коробочкой со спорами; заростком.)
- Какое поколение преобладает в жизненном цикле папоротников? (Бесполое — спорофит; половое — гаметофит.)
- Что служит ограничивающим фактором в цикле развития папоротников? (Воздух, вода, температура.)
Ø Деятельность учащихся
Подписывают рисунок.
Предполагаемые правильные ответы:
- Гаметофит.
- Заростком.
- Травянистым растением с корневищем, стеблем и листьями.
- Спорофит.
- Вода.
- Рефлексия
Ø Деятельность учителя
Создает рефлексивную ситуацию, чтобы получить качественную оценку работы на уроке каждого учащегося и всего класса в целом.
Оценивает активность учащихся на уроке, благодарит за работу. Предлагает учащимся заполнить анкету для рефлексии. Анкета не подписывается.
Пример анкеты для рефлексии:
- На уроке я работал … активно / пассивно
- Своей работой на уроке я … доволен / не доволен
- Мне хочется, чтобы …
Ø Деятельность учащихся
Оценивают результаты собственной работы на уроке. Анонимно заполняют анкету и отдают учителю.
- Домашнее задание
Ø Деятельность учителя
Предлагает выполнить следующие задания. Изучить § 42 учебного пособия. Ответить на вопросы после параграфа.
Сообщает учащимся, что на следующем уроке речь пойдет о многообразии папоротников, поэтому необходимо подготовить сообщения по темам: “Папоротники — комнатные растения”, “Лекарственные папоротники”, “Папоротники — пищевые растения”, “Папоротники, занесенные в Красную книгу”.
Ø Деятельность учащихся
Записывают домашнее задание. Готовят дома сообщения.
краткая схема посадки и пересадки
Папоротники — древнейшие растения, живущие на нашей планете. На Земле их насчитывают около 11.000 видов. Благодаря своим оригинальным листьям, они давно заслужили внимание цветоводов и стали использоваться, как домашние растения в качестве украшения интерьера и помощников в очищении и увлажнении воздуха.
Папоротник как домашнее и садовое растение – 2 вида
В природе существует два известных нам вида папоротников:
- спорофиты — бесполые папоротники;
- гаметофиты — половые папоротники (мужского и женского пола).
Представители первого вида произрастают в субтропиках, их листья сначала свернуты в спираль и постепенно с развитием растения разворачиваются. Именно спорофиты содержатся в качестве комнатных растений в домах и квартирах цветоводов. Форма и размер листьев у каждого сорта папоротника отличается от других. Растут листья весьма медленно. Листовая почка развивается под землей в течение нескольких лет, прежде чем показаться на свет в виде улиткообразной пластины.
Для комнатного разведения подходят следующие виды папоротников:
- Адиантум (Adiantum) – изящное растение с перистыми листьями, расположенными на тонких темно-коричневых черешках. Он весьма быстро растет, неприхотлив, неплохо переносит затемненные места.
- Асплениум (Asplenium) привлекает внимание блестящими цельными листьями светло-зеленого цвета, по центру каждого из которых проходит выразительная темная жилка.
- Нефролепис (Nephrolepis) – неприхотливое растение с ажурными воздушными длинными листьями светло-зеленого цвета.
Поливать папоротник в домашних условиях следует по мере просыхания верхней части грунта, это растение любит воду, в поливе заключается практически весь уход за ним. Пересадка комнатного папоротника проводится только весной, в случае если предыдущий горшок стал ему тесен. Одним из самых популярных садовых папоротников является страусник лесной обыкновенный, называемый также черный папоротник. Он спокойно переносит морозы до -20 градусов, быстро растет в саду. Часто его используют, как декоративное растение на даче, он предпочитает тенистые места, которые обычно пустуют.
Размножение спорами – подготовка почвы, техника проращивания
Процесс размножения папоротников спорами заключается в самом сборе созревших спор, их хранении с целью дозревания и непосредственно высадке в грунт. Споры папоротника расположены в сорусах, которые приобретают бурый оттенок при созревании в них спор. Срезать споры необходимо вместе с частью листа, а затем хранить в плотно запечатанном пакете из бумаги до полного дозревания. Созревшие и готовые к высадке в почву споры представляют собой порошкообразное вещество коричневого цвета.
Для посева спор папоротника приготовьте одну из ниже представленных смесей почвы:
- песок и бурый торф – 1:3;
- песок, листовую землю и бурый торф – 1:1:2;
- песок и листовую землю – 1:3;
- готовый грунт для фиалок.
Смесь нужно будет очистить от возможно имеющихся крупных частиц и прогреть в духовой печи, для того чтобы уничтожить вредителей и семена сорных трав. В качестве емкости для высаживания спор папоротника используйте неглубокий контейнер, толщина слоя почвы в нем должна быть около 5 см. На хорошо утрамбованную и политую заранее почву равномерно распределяют споры, не присыпая их сверху. Контейнер накрывают стеклом и ставят в светлом, но защищенном от солнечных лучей месте, при комнатной температуре.
Время прорастания спор напрямую зависит от вида папоротника, это может быть 3 недели, а может занять и год. На протяжении всего времени всхода спор поддерживайте влажность грунта, в котором они находятся, только при помощи увлажнения поддона. Время от времени открывайте стекло, чтобы пустить к спорам кислород. Проросшие споры папоротника выглядят, как мох, покрывающий всю поверхность почвы. Пророщенные папоротники в самом начале не имеют корней, они начинают появляться только спустя 4-6 недель.
Покрывное стекло убираем постепенно, начиная раскрывать растения на пару часов в сутки, и каждый раз увеличивая период времени. Как только вы приучили папоротник к отсутствию парникового эффекта, начинайте опрыскивать ростки водой. 2 раза в неделю можно растворять в воде Эпин или другие микроэлементы и опрыскивать растения курсами по 2 недели в месяц. Развитое растение у вас сформируется через полгода после всходов, пересаживать папоротник в индивидуальный горшок можно, как только начнет развиваться корневая система.
Этот метод используется самыми терпеливыми цветоводами. За рождением новых представителей папоротниковых очень интересно наблюдать, стадии их развития отличаются от роста большинства других растений, но уже изначально следует понимать, что этот процесс займет много времени и будет требовать от вас заботы и поддержания комфортных климатических условий окружающей среды.
Вегетативное размножение – деление корней, почкование
Куда проще размножить папоротник вегетативным путем. Способом деления корней размножаются садовые разновидности папоротника, несколько видов комнатных, такие как: листовики, адиантумы и пеллеи. Таким образом, можно размножить папоротник в процессе пересадки, весной.
Заметьте, для размножения подходят только крепкие взрослые растения, которые имеют более одной точки роста.
Деление проводят в прохладном помещении – корневую систему аккуратно разрезают, отделяя розетки друг от друга, а затем высаживают каждый отросток в грунт и обильно поливают. При правильном проведении процедуры приживаемость растений очень высока, так как каждый отросток имеет уже развитые корни, способные поглощать из почвы необходимые для роста вещества и микроэлементы, а также воду. В течение некоторого времени корневая система восстановит поврежденные участки и начнет развитие новых ответвлений.
Есть некоторые виды папоротников, которые размножаются посредством почек на черешках. Это, например, пузырник луковиценосный. В таких случаях почки пересаживают в тепличные условия, где поддерживается постоянная влажность. После того, как почки укоренятся, их можно пересадить в отдельный небольшого размера горшочек вместе с частью привычного им грунта. Далее правила ухода будут те же, что и для молодых папоротников, выращенных любым другим способом.
Как видно, папоротник сравнительно некапризное растение, важно лишь учитывать его тропическое происхождение и создать условия, идентичные тем, к которым он привык. Взамен вы получите прекрасного помощника в озеленении вашего домашнего интерьера или садового участка. Специалисты рекомендуют ставить горшок с папоротником в детских комнатах, неподалеку от письменного стола ребенка – это поспособствует концентрации внимания и усидчивости, благодаря вышеупомянутой способности растения очищать воздушное пространство и насыщать его кислородом.
Значениедля потенциала колонизации JSTOR
Abstract• Предпосылки и цели Изменения окружающей среды, опосредованные человеком, усиливают давление отбора на способность растений колонизировать новые области. Фрагментация среды обитания в сочетании с изменением климата в целом вынуждает виды колонизировать районы на больших расстояниях. Системы спаривания и генетическая нагрузка являются важными детерминантами создания и долгосрочного выживания новых популяций. Здесь система спаривания Asplenium scolopendnum, диплоидного гомоспористого вида папоротников, исследуется в связи с процессами колонизации.• Методы. Эксперимент в общей среде был проведен с 13 парами спорофитов, каждая из которых была выращена на разных участках. Вместе они составляют по крайней мере девять различных генотипов, представляющих, по оценкам, прибл. 95% нечастных внутривидовых генетических вариаций в Европе. Продукцию спорофитов регистрировали для гаметофитов, происходящих от каждого родительского спорофита. Гаметофиты выращивали in vitro тремя различными способами: (I) изолированно, (II) с гаметофитом из другого спорофита в пределах одного и того же сайта или (III) с партнером из другого сайта.• Ключевые результаты Продукция спорофитов была наивысшей при межсайтовых скрещиваниях (III), промежуточной при внутрисайтовых скрещиваниях (II) и была наименьшей при изолированных гаметофитах (I), что явно указывает на инбридинговую депрессию. Однако внутригаметофитное самоопыление наблюдалось у большинства протестированных генотипов (восемь из девяти). • Выводы. Результаты предполагают смешанную систему спаривания у A. scolopendnum с ауткроссингом, когда это возможно, и периодическим самоопылением, когда это необходимо. Периодическое внутриигаметофитное самоопыление способствует успешному заселению новых участков из одной споры.Образовавшийся спорофит, который будет полностью гомозиготным, со временем будет выделять большое количество спор. Каждый год это создает ложе гаметофитов в непосредственной близости от родителей. Любая неродственная спора, которая появляется, затем выборочно воспроизводится и вносит свои гены в новую популяцию. Таким образом, в то время как самоопыление способствует первоначальному успеху колонизации, инбридинговая депрессия способствует генетически разнообразным популяциям посредством ауткроссинга. Результаты являются дополнительным доказательством против слишком простого дихотомического разделения видов папоротников на самоопыление или ауткроссинг.
Информация об издателеOxford University Press — это отделение Оксфордского университета. Издание во всем мире способствует достижению цели университета в области исследований, стипендий и образования. OUP — крупнейшая в мире университетская пресса с самым широким глобальным присутствием. В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5500 сотрудников по всему миру. Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные работы по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы.
Системы разведения гомоспористых папоротников. A, Интрагаметофитное самоопыление, …
Контекст 1
… с образованием гемм у некоторых таксонов (например, многих Vittariaceae) продолжительность жизни может быть значительно увеличена. Например, колонии гаметофитов Hymenophyllaceae и Vittariaceae в восточной части США отслеживаются более 50 лет и могут быть реликтами доплейстоценового расширения ареала рассматриваемых видов (Farrar 1967).Большинство этих видов продуцируют ветвящиеся геммоносные гаметофиты. Еще одно важное преимущество неопределенного роста заключается в том, что долгоживущие гаметофиты могут достигать размеров в несколько квадратных сантиметров и производить десятки спорофитов в пространстве и времени (Фаррар и др., 2008; рис. 4), в отличие от модели одного гаметофита и одного спорофита, изображенной на учебники. Watkins et al. (2007 b) обнаружили, что основной причиной смертности наземных видов было нарушение. Фактически, ни один наземный вид (отмеченный как гаметофит и последующий пополнение спорофитов) не выжил достаточно долго, чтобы произвести плодородный спорофит.Это и другие исследования (Peck et al. 1990) указывают на то, что наземные виды при укоренении зависят от нарушений. Немногие виды способны расти на или под опавшей листвой, и необходимо вмешательство, чтобы обнажить нижележащую минеральную почву. Виды в этих средах обитания проходят r-отбор, и им необходимо быстро пройти свой жизненный цикл до того, как их заменит подстилка или конкурирующие растения. Покров, напротив, может быть менее подвержен нарушениям (Watkins et al. 2007 b), и такая стабильность будет способствовать появлению долгоживущих гаметофитов, которые могут достигать больших размеров (рис.4). Гипотеза о том, что повышение стабильности среды обитания может способствовать долголетию, не нова (Grime 1977). Тем не менее, остается вопрос, почему папоротники, растущие в эпифитных средах обитания, получают выгоду от увеличения продолжительности жизни гаметофитов? Чтобы понять это, нужно отказаться от простого бессмысленного хрестоматийного представления о самоопыляющемся гаметофите папоротника (Campbell 2004; Freeman 2011). Студент, полагающийся на текущее описание жизненного цикла папоротника в учебниках, быстро придет к выводу, что распространение спор папоротника эквивалентно миграции.Путем внутригаметофитного самоопыления (рис. 5 A) одна спора может дать начало спорофиту, который продолжает жизненный цикл. Любой спорофит, продуцируемый в рамках этой системы, будет гомозиготным по всем локусам (сперматозоид и яйцеклетка происходят от одного и того же мейотического события). Любой спорофитно-рецессивный летальный или иным образом вредный ген может предотвратить образование спорофита или отрицательно повлиять на него. Тем не менее, ранняя работа Клековски (1979) по выборочному набору видов предполагала, что такое самоопыление может быть обычным явлением среди папоротников, особенно с подземными гаметофитами (Soltis and Soltis 1986).Однако данные по изоферментам показывают, что межгаметофитное скрещивание (рис. 5 C) является правилом для диплоидных гомоспористых папоротников с надземными гаметофитами (Haufler and Soltis 1984; Soltis and Soltis 1987, 1990; Chiou et al. 1998, 2002). Klekowski (1979) действительно предполагает, что повышенная экологическая стабильность ведет к увеличению продолжительности жизни гаметофитов и межгаметофитному спариванию, что является явно эпифитной стратегией. Один из способов уменьшить трудности получения гаметангий из отдельных гаметофитов вместе — это произвести долгоживущий гаметофит, который может «ждать», пока поблизости не приземлится другой подходящий генотип.Фаррар и др. (2008) подробно обсудили вариации в форме гаметофита. Уместным для нашего обсуждения является то, что многие эпифитные папоротники соединились с медленнорастущими многолетними гаметофитами в форме лент и лент. В отличие от короткоживущих сердцевидных гаметофитов наземных видов, эпифитные гаметофиты могут существовать дольше и лучше конкурировать в матах мохообразных, встречающихся во многих зрелых эпифитных местообитаниях. Неопределенная трехмерная форма эпифитных гаметофитов папоротников (рис.4) позволяет им исследовать эти среды обитания в поисках микросайтов, где они могут сохраняться и ждать благоприятных условий для образования спорофитов (Dassler and Farrar 1997; Farrar et al.2008 г.). Эта долгоживущая стратегия «банка гаметофитов» невозможна для большинства наземных видов, которые при укоренении полагаются на нарушение порядка (Watkins et al. 2007 b). Дополнительной стратегией, разработанной во многих эпифитных клонах, является бесполое размножение с помощью диспергируемых гемм (рис. 4 H-4 L; Dassler and Farrar 1997). Виды с драгоценными гаметофитами обладают дополнительным преимуществом распространения пропагул, полученных от гаметофитов-основателей, на новые микросайты. И долгоживущие, и драгоценные гаметофиты могут иметь решающее значение для укоренения папоротников за счет распространения на большие расстояния, когда вероятность объединения двух спор одновременно крайне маловероятна.Важным компонентом увеличения продолжительности жизни гаметофитов является способность гаметофитов сохраняться в областях, где выживание спорофитов в настоящее время невозможно, но может стать возможным с изменением окружающей среды. Известно, что происходит самоопыление, и первые виды папоротников являются классическими примерами этого, но ауткроссинг преобладает у большинства видов (Tryon 1970; Crist and Farrar 1983; Ranker et al. 1994; Moran and Smith 2001). Папоротники разработали ряд разнообразных химических систем определения пола (т. Е., антеридиогены [Schneller 2008] и сложные системы разведения для поддержания гетерозиготности). Теперь мы знаем, что гаметофиты большинства видов функционально не двуполые, поддерживают разумный уровень генетического разнообразия и несут сильно различающийся объем генетической нагрузки (Ranker and Geiger 2008). Гаметофит, нашедший безопасное место для созревания, должен подождать, пока другой гаметофит (либо от того же спорофита, возможно, либо от другого растения) приземлится достаточно близко для передачи сперматозоидов, что является почти неразрешимой проблемой на любом расстоянии от спорофита.Если склонность к самосознанию редка, а долгосрочные последствия такого спаривания отрицательны (Ранкер и Гейгер, 2008), то можно ожидать, что у островных популяций сократится генетическое разнообразие. К сожалению, такие данные недоступны. Однако мы можем сделать некоторые выводы из флористического анализа. В сравнительном обзоре островных и континентальных флор Фаррар и др. (2008) обнаружили, что, хотя общее количество эпифитов одинаково в обоих регионах, островные флоры увеличили изобилие эпифитных видов с драгоценными гаметофитами.Может случиться так, что полог ослабляет давление инбридинга и способствует более быстрому видообразованию — гипотеза, согласующаяся с паническим бегством птеридофитов мелового периода в полог. Одним из основных факторов, с которыми ежедневно сталкиваются гаметофиты, является доступность воды; это естественно усиливается в таксонах, которые живут годами, особенно в эпифитных местообитаниях. Watkins et al. (2007 a) исследовали степень устойчивости к высыханию гаметофитов ряда тропических эпифитных и наземных видов.Это исследование выявило удивительную степень устойчивости к высыханию у гаметофитов всех видов, но особенно у эпифитных таксонов. Например, после того, как гаметофиты подверглись действию и регидратации из-за дефицита давления пара À 94 МПа (чрезвычайно сухой воздух для любого растения) в течение 24 часов, все виды показали более 50% …
Контекст 2
… при производстве гемм в некоторых таксонах (например, во многих Vittariaceae) продолжительность жизни может быть значительно увеличена.Например, колонии гаметофитов Hymenophyllaceae и Vittariaceae в восточной части США отслеживаются более 50 лет и могут быть реликтами доплейстоценового расширения ареала рассматриваемых видов (Farrar 1967). Большинство этих видов продуцируют ветвящиеся геммоносные гаметофиты. Еще одно важное преимущество неопределенного роста заключается в том, что долгоживущие гаметофиты могут достигать размеров в несколько квадратных сантиметров и производить десятки спорофитов в пространстве и времени (Farrar et al., 2008; рис.4), в отличие от одного гаметофита в учебниках — одна модель спорофита. Watkins et al. (2007 b) обнаружили, что основной причиной смертности наземных видов было нарушение. Фактически, ни один наземный вид (отмеченный как гаметофит и последующий пополнение спорофитов) не выжил достаточно долго, чтобы произвести плодородный спорофит. Это и другие исследования (Peck et al. 1990) указывают на то, что наземные виды при укоренении зависят от нарушений. Немногие виды способны расти на или под опавшей листвой, и необходимо вмешательство, чтобы обнажить нижележащую минеральную почву.Виды в этих средах обитания проходят r-отбор, и им необходимо быстро пройти свой жизненный цикл до того, как их заменит подстилка или конкурирующие растения. Покров, напротив, может быть менее подвержен нарушениям (Watkins et al. 2007 b), и такая стабильность будет способствовать появлению долгоживущих гаметофитов, которые могут достигать больших размеров (рис. 4). Гипотеза о том, что повышение стабильности среды обитания может способствовать долголетию, не нова (Grime 1977). Тем не менее, остается вопрос, почему папоротники, растущие в эпифитных средах обитания, получают выгоду от увеличения продолжительности жизни гаметофитов? Чтобы понять это, нужно отказаться от простого бессмысленного хрестоматийного представления о самоопыляющемся гаметофите папоротника (Campbell 2004; Freeman 2011).Студент, полагающийся на текущее описание жизненного цикла папоротника в учебниках, быстро придет к выводу, что распространение спор папоротника эквивалентно миграции. Путем внутригаметофитного самоопыления (рис. 5 A) одна спора может дать начало спорофиту, который продолжает жизненный цикл. Любой спорофит, продуцируемый в рамках этой системы, будет гомозиготным по всем локусам (сперматозоид и яйцеклетка происходят от одного и того же мейотического события). Любой спорофитно-рецессивный летальный или иным образом вредный ген может предотвратить образование спорофита или отрицательно повлиять на него.Тем не менее, ранняя работа Клековски (1979) по выборочному набору видов предполагала, что такое самоопыление может быть обычным явлением среди папоротников, особенно с подземными гаметофитами (Soltis and Soltis 1986). Однако данные по изоферментам показывают, что межгаметофитное скрещивание (рис. 5 C) является правилом для диплоидных гомоспористых папоротников с надземными гаметофитами (Haufler and Soltis 1984; Soltis and Soltis 1987, 1990; Chiou et al. 1998, 2002). Klekowski (1979) действительно предполагает, что повышенная экологическая стабильность ведет к увеличению продолжительности жизни гаметофитов и межгаметофитному спариванию, что является явно эпифитной стратегией.Один из способов уменьшить трудности получения гаметангий из отдельных гаметофитов вместе — это произвести долгоживущий гаметофит, который может «ждать», пока поблизости не приземлится другой подходящий генотип. Фаррар и др. (2008) подробно обсудили вариации в форме гаметофита. Уместным для нашего обсуждения является то, что многие эпифитные папоротники соединились с медленнорастущими многолетними гаметофитами в форме лент и лент. В отличие от короткоживущих сердцевидных гаметофитов наземных видов, эпифитные гаметофиты могут существовать дольше и лучше конкурировать в матах мохообразных, встречающихся во многих зрелых эпифитных местообитаниях.Неопределенная трехмерная форма эпифитных гаметофитов папоротников (рис. 4) позволяет им исследовать эти среды обитания в поисках микросайтов, где они могут сохраняться и ждать благоприятных условий для образования спорофитов (Dassler and Farrar 1997; Farrar et al. 2008). Эта долгоживущая стратегия «банка гаметофитов» невозможна для большинства наземных видов, которые при укоренении полагаются на нарушение порядка (Watkins et al. 2007 b). Дополнительная стратегия, разработанная во многих эпифитных линиях, — это бесполое размножение с помощью диспергируемых гемм (рис.4 H –4 л; Дасслер и Фаррар 1997). Виды с драгоценными гаметофитами обладают дополнительным преимуществом распространения пропагул, полученных от гаметофитов-основателей, на новые микросайты. И долгоживущие, и драгоценные гаметофиты могут иметь решающее значение для укоренения папоротников за счет распространения на большие расстояния, когда вероятность объединения двух спор одновременно крайне маловероятна. Важным компонентом увеличения продолжительности жизни гаметофитов является способность гаметофитов сохраняться в областях, где выживание спорофитов в настоящее время невозможно, но может стать возможным с изменением окружающей среды.Известно, что происходит самоопыление, и первые виды папоротников являются классическими примерами этого, но ауткроссинг преобладает у большинства видов (Tryon 1970; Crist and Farrar 1983; Ranker et al. 1994; Moran and Smith 2001). Папоротники разработали ряд разнообразных химических систем определения пола (например, антеридиогены [Schneller 2008] и сложные системы размножения для поддержания гетерозиготности). Теперь мы знаем, что гаметофиты большинства видов функционально не двуполые, поддерживают разумный уровень генетического разнообразия и несут сильно различающийся объем генетической нагрузки (Ranker and Geiger 2008).Гаметофит, нашедший безопасное место для созревания, должен подождать, пока другой гаметофит (либо от того же спорофита, возможно, либо от другого растения) приземлится достаточно близко для передачи сперматозоидов, что является почти неразрешимой проблемой на любом расстоянии от спорофита. Если склонность к самосознанию редка, а долгосрочные последствия такого спаривания отрицательны (Ранкер и Гейгер, 2008), то можно ожидать, что у островных популяций сократится генетическое разнообразие. К сожалению, такие данные недоступны.Однако мы можем сделать некоторые выводы из флористического анализа. В сравнительном обзоре островных и континентальных флор Фаррар и др. (2008) обнаружили, что, хотя общее количество эпифитов одинаково в обоих регионах, островные флоры увеличили изобилие эпифитных видов с драгоценными гаметофитами. Может случиться так, что полог ослабляет давление инбридинга и способствует более быстрому видообразованию — гипотеза, согласующаяся с паническим бегством птеридофитов мелового периода в полог. Одним из основных факторов, с которыми ежедневно сталкиваются гаметофиты, является доступность воды; это естественно усиливается в таксонах, которые живут годами, особенно в эпифитных местообитаниях.Watkins et al. (2007 a) исследовали степень устойчивости к высыханию гаметофитов ряда тропических эпифитных и наземных видов. Это исследование выявило удивительную степень устойчивости к высыханию у гаметофитов всех видов, но особенно у эпифитных таксонов. Например, после того, как гаметофиты подверглись действию и регидратации из-за дефицита давления пара À 94 МПа (чрезвычайно сухой воздух для любого растения) в течение 24 часов, все виды показали более 50% …
Смешанная система спаривания в папоротник Asplenium scolopendrium: значение для потенциала колонизации | Летопись ботаники
5″ data-legacy-id=»s1″> ВВЕДЕНИЕ Экосистемы все больше подвергаются стрессу из-за антропогенных изменений окружающей среды, и выживание видов становится все более и более зависимым от способности колонизировать новые районы (Cain et al., 2000; Thomas et al. , 2004). Фрагментация среды обитания увеличивает среднее расстояние между участками подходящей среды обитания, в то время как в более крупном пространственном масштабе изменение климата одновременно вынуждает виды менять свои естественные ареалы (Svenning and Condit, 2008; Svenning et al. , 2008, 2009). Помимо проблем расселения в новые районы и местного обоснования, долгосрочный успех новых колоний зависит от репродуктивного успеха их колонистов (например, Soons and Heil, 2002).Увеличение размера местной популяции при первоначальной колонизации необходимо для снижения риска местного исчезновения, в то время как увеличение генетического разнообразия часто также важно в этом отношении (например, Spielman et al. , 2004; Crawford and Whitney, 2010).
Вероятность найти партнера по достижении половой зрелости быстро снижается по мере того, как отростки расходятся дальше от родителей (Peck et al. , 1990; Nathan, 2001). Поэтому колонизирующие растения больше полагаются на самооплодотворение и будут испытывать длительные периоды спаривания между родственниками.У многих видов самоопыление или инбридинг из-за небольшого размера популяции может привести к снижению приспособленности инбредного потомства к беспородному, то есть к депрессии инбредного скрещивания (Crnokrak and Roff, 1999; Keller and Waller, 2002, но см. , 2009). Экспериментальные исследования селекции предоставляют информацию о том, в какой степени инбридинговая депрессия повлияет на индивидуальную приспособленность, что позволяет оценить генетические риски, которые изменения окружающей среды представляют для выживания.
Папоротники представляют собой относительно малоизученную группу в Царстве растений, и для этой группы существует общий недостаток знаний о процессах колонизации и роли в них систем спаривания (Haufler, 2009).Папоротники отличаются от семенных растений тем, что распространение происходит через гаплоидные споры, а их жизненный цикл включает независимое поколение гаметофитов. Обычно считается, что гаметофиты в современных папоротниках недолговечны, хотя экспериментальных данных мало (но см. Sato, 1982; Peck et al. , 1990), и существуют заметные исключения (например, Osmundaceae и граммитиды у Polypodiaceae). Спорофиты являются диплоидными (или полиплоидными производными в некоторых таксонах), а взрослые особи производят несколько миллионов спор каждый год.По сравнению с семенами, из-за своего небольшого размера споры, как правило, разносятся ветром гораздо дальше. Однако, как и у семенных растений, большинство пропагул приземляются в непосредственной близости от родителя (Peck et al. , 1990). Дистанция распространения мужских гамет обычно очень ограничена, поскольку сперматозоиды требуют транспортировки в воде. Хотя папоротники производят большое количество спор и разносятся ветром относительно легко, успешное спаривание после распространения на большие расстояния [определено как> 1 км Peck et al. (1990)], требующий одновременного развития двух гаметофитов в пределах нескольких сантиметров, считается маловероятным событием (Peck et al. , 1990; Flinn, 2006). Однако частотное распределение папоротников на расстояние распространения до сих пор в значительной степени неизвестно, и поэтому неясно, как часто такие события действительно происходят.
Односпоровые папоротники образуют споры одного типа, которые могут дать начало гаметофитам, которые потенциально могут стать двуполыми. Самооплодотворение гаплоидного гаметофита, т.е. внутригаметофитное самоопыление, представляет собой крайний случай инбридинга и приводит к полностью гомозиготным спорофитам (Vogel et al. , 1999 b ). Таким образом, при отсутствии генетической нагрузки для успешной колонизации новых участков требуется только одна спора (Lloyd, 1974). Помимо гаметофитного самоопыления intra , аналогов которому нет у семенных растений, у папоротников возможно самоопыление inter гаметофита, когда объединяются гаметы двух гаметофитов, происходящих от одного и того же спорофита; это эквивалентно самоопылению семенных растений.Ауткроссинг происходит за счет объединения гамет двух гаметофитов, происходящих от разных спорофитов, хотя это все еще может эффективно представлять собой инбридинг, когда родительские спорофиты тесно связаны (т. Е. Межродительский инбридинг). На основании экспериментов по разведению и анализа генетической изменчивости гомоспористые папоротники в прошлом классифицировались либо как преимущественно ауткроссинговые, либо как самоопыляющиеся виды (Soltis and Soltis, 1992). По сравнению с диплоидами, полиплоидные виды папоротников, как правило, менее восприимчивы к депрессии инбридинга и выдерживают более высокие показатели самоопыления (Masuyama, 1979; Soltis and Soltis, 2000).
В этой статье представлена система размножения Asplenium scolopendrium (Aspleniaceae), диплоидных (2 n = 72) гомоспористых видов папоротников (Tutin et al. , 1993), которые встречаются по всей Западной Европе (Jalas and Suominen , 1988). Наблюдения за онтогенезом гаметофитов для некоторых шотландских популяций предложили стратегию ауткроссинга для A. scolopendrium (Pangua et al. , 1994). Этот вид успешно колонизировал ряд лесов на недавно высаженном дамбой (1942 г.) польдере в Нидерландах (Bremer and Jongejans 2010).По оценкам, численность популяции спорофитов в 2002 году составляла> 13 000, это один из самых распространенных видов папоротников в одном из этих лесов — «Куиндербос» (Bremer, 2007). Мы предположили, что ауткроссинг у этого вида должен быть более успешным, чем самоопыление. Во-первых, потому что, по крайней мере, в Европе A. scolopendrium является диплоидным таксоном, а во-вторых, потому что его онтогенез гаметофитов предполагает стратегию ауткроссинга (Pangua et al. , 1994). Кроме того, учитывая очевидный успех колонизации голландских польдеров, а также всей Европы (Vogel et al., 1999, b ), его потенциал для внутригаметофитного самоопыления представлял интерес.
2″ data-legacy-id=»s2a»> Район исследования и полевой отбор
Экосистемы все больше подвергаются стрессу из-за антропогенных изменений окружающей среды, и выживание видов становится все более и более зависимым от способности колонизировать новые районы (Cain et al., 2000; Thomas et al. , 2004). Фрагментация среды обитания увеличивает среднее расстояние между участками подходящей среды обитания, в то время как в более крупном пространственном масштабе изменение климата одновременно вынуждает виды менять свои естественные ареалы (Svenning and Condit, 2008; Svenning et al. , 2008, 2009). Помимо проблем расселения в новые районы и местного обоснования, долгосрочный успех новых колоний зависит от репродуктивного успеха их колонистов (например, Soons and Heil, 2002).Увеличение размера местной популяции при первоначальной колонизации необходимо для снижения риска местного исчезновения, в то время как увеличение генетического разнообразия часто также важно в этом отношении (например, Spielman et al. , 2004; Crawford and Whitney, 2010).
Вероятность найти партнера по достижении половой зрелости быстро снижается по мере того, как отростки расходятся дальше от родителей (Peck et al. , 1990; Nathan, 2001). Поэтому колонизирующие растения больше полагаются на самооплодотворение и будут испытывать длительные периоды спаривания между родственниками.У многих видов самоопыление или инбридинг из-за небольшого размера популяции может привести к снижению приспособленности инбредного потомства к беспородному, то есть к депрессии инбредного скрещивания (Crnokrak and Roff, 1999; Keller and Waller, 2002, но см. , 2009). Экспериментальные исследования селекции предоставляют информацию о том, в какой степени инбридинговая депрессия повлияет на индивидуальную приспособленность, что позволяет оценить генетические риски, которые изменения окружающей среды представляют для выживания.
Папоротники представляют собой относительно малоизученную группу в Царстве растений, и для этой группы существует общий недостаток знаний о процессах колонизации и роли в них систем спаривания (Haufler, 2009).Папоротники отличаются от семенных растений тем, что распространение происходит через гаплоидные споры, а их жизненный цикл включает независимое поколение гаметофитов. Обычно считается, что гаметофиты в современных папоротниках недолговечны, хотя экспериментальных данных мало (но см. Sato, 1982; Peck et al. , 1990), и существуют заметные исключения (например, Osmundaceae и граммитиды у Polypodiaceae). Спорофиты являются диплоидными (или полиплоидными производными в некоторых таксонах), а взрослые особи производят несколько миллионов спор каждый год.По сравнению с семенами, из-за своего небольшого размера споры, как правило, разносятся ветром гораздо дальше. Однако, как и у семенных растений, большинство пропагул приземляются в непосредственной близости от родителя (Peck et al. , 1990). Дистанция распространения мужских гамет обычно очень ограничена, поскольку сперматозоиды требуют транспортировки в воде. Хотя папоротники производят большое количество спор и разносятся ветром относительно легко, успешное спаривание после распространения на большие расстояния [определено как> 1 км Peck et al. (1990)], требующий одновременного развития двух гаметофитов в пределах нескольких сантиметров, считается маловероятным событием (Peck et al. , 1990; Flinn, 2006). Однако частотное распределение папоротников на расстояние распространения до сих пор в значительной степени неизвестно, и поэтому неясно, как часто такие события действительно происходят.
Односпоровые папоротники образуют споры одного типа, которые могут дать начало гаметофитам, которые потенциально могут стать двуполыми. Самооплодотворение гаплоидного гаметофита, т.е. внутригаметофитное самоопыление, представляет собой крайний случай инбридинга и приводит к полностью гомозиготным спорофитам (Vogel et al. , 1999 b ). Таким образом, при отсутствии генетической нагрузки для успешной колонизации новых участков требуется только одна спора (Lloyd, 1974). Помимо гаметофитного самоопыления intra , аналогов которому нет у семенных растений, у папоротников возможно самоопыление inter гаметофита, когда объединяются гаметы двух гаметофитов, происходящих от одного и того же спорофита; это эквивалентно самоопылению семенных растений.Ауткроссинг происходит за счет объединения гамет двух гаметофитов, происходящих от разных спорофитов, хотя это все еще может эффективно представлять собой инбридинг, когда родительские спорофиты тесно связаны (т. Е. Межродительский инбридинг). На основании экспериментов по разведению и анализа генетической изменчивости гомоспористые папоротники в прошлом классифицировались либо как преимущественно ауткроссинговые, либо как самоопыляющиеся виды (Soltis and Soltis, 1992). По сравнению с диплоидами, полиплоидные виды папоротников, как правило, менее восприимчивы к депрессии инбридинга и выдерживают более высокие показатели самоопыления (Masuyama, 1979; Soltis and Soltis, 2000).
В этой статье представлена система размножения Asplenium scolopendrium (Aspleniaceae), диплоидных (2 n = 72) гомоспористых видов папоротников (Tutin et al. , 1993), которые встречаются по всей Западной Европе (Jalas and Suominen , 1988). Наблюдения за онтогенезом гаметофитов для некоторых шотландских популяций предложили стратегию ауткроссинга для A. scolopendrium (Pangua et al. , 1994). Этот вид успешно колонизировал ряд лесов на недавно высаженном дамбой (1942 г.) польдере в Нидерландах (Bremer and Jongejans 2010).По оценкам, численность популяции спорофитов в 2002 году составляла> 13 000, это один из самых распространенных видов папоротников в одном из этих лесов — «Куиндербос» (Bremer, 2007). Мы предположили, что ауткроссинг у этого вида должен быть более успешным, чем самоопыление. Во-первых, потому что, по крайней мере, в Европе A. scolopendrium является диплоидным таксоном, а во-вторых, потому что его онтогенез гаметофитов предполагает стратегию ауткроссинга (Pangua et al. , 1994). Кроме того, учитывая очевидный успех колонизации голландских польдеров, а также всей Европы (Vogel et al., 1999, b ), его потенциал для внутригаметофитного самоопыления представлял интерес.
Куиндербос (52,8 ° с.ш., 5,8 ° в.д.), Нидерланды, представляет собой недавно высаженный лес (1949–1954 гг.) На земле, которая была восстановлена у бывшего морского залива «Зуйдерзее» в 1942 году. Растительность представлена равномерно возрастными насаждениями деревьев ( Fagus sylvatica , Fraxinus excelsior , Picea abies , P.sitchensis и Quercus robur ), а также различные виды подроста. К последним относятся в Нидерландах исключительно большое количество папоротников (Bremer, 1980, 2007) и мохообразных (Bremer and Ott, 1990). Почва состоит из слоя известковых песков (pH ≥ 7 · 2) поверх торфа. Верхний слой песка пересекается многочисленными дренажными канавами, которые хорошо врезаются в нижний слой торфа (Bremer, 2007). Это создало прекрасную среду для папоротников, и на местном уровне с этими желобами тесно связан Asplenium scolopendrium (Bremer, 1980, 2007).Подробные данные инвентаризации были собраны в этом районе за последние 30 лет (Bremer, 2007), что позволяет разграничить отдельные места колонизации. Места колонизации определялись как группы людей, занимающие одну и ту же траншею, с распределением особей в пределах участков не более чем на несколько метров.
Целые плодородные листья были собраны на 13 участках A. scolopendrium по всей территории Куиндербоса, взяв при этом листья двух спороносных растений на каждом участке.Отобранные спорофиты анализировали на аллозимную изменчивость в 11 локусах, следуя стандартным методам (см. Vogel et al. , 1999, и ). Ваучеры с исследованных мест колонизации были депонированы в Лондонском музее естественной истории (BM). Были выделены следующие 11 ферментных систем: 6-фосфоглюконатдегидрогеназа (6-PGD, EC 1 · 1 · 1 · 44), аспартатаминотрансфераза (AAT, EC 2 · 6 · 1 · 1), кислая фосфатаза (ACP, EC 3 · 1 · 3 · 2), глутаматдегидрогеназа (GDH, EC 1 · 4 · 1 · 2), изоцитратдегидрогеназа (IDH, EC 1 · 1 · 1 · 42), лейцинаминопептидаза (LAP, EC 3 · 4 · 11 · 1 ), малатдегидрогеназа (MDH, EC 1 · 1 · 1 · 37), фосфоглюкозоизомераза (PGI, EC 5 · 3 · 1 · 9), шикиматдегидрогеназа (SkDH, EC 1 · 1 · 1 · 25), триозофосфат изомераза (TPI, EC 5 · 3 · 1 · 1) и utp-глюкозопирофосфорилаза (UGPP, EC 2 · 7 · 9).Образцы из семи популяций по всей Европе (Англия, n = 8 и 24; Франция, 30; Германия, 10; Греция, 14; Шотландия, 21; Уэльс, 32) также были проанализированы на предмет аллозимной изменчивости в этих локусах для оценки генетического изменчивость включена в экспериментальную часть данного исследования (см. ниже). Эти данные являются частью более крупного популяционного генетического исследования этого вида (J. C. Vogel et al. , неопубликованные данные).
Генотипы очаговых особей
. Локус
. 1
. 2
. 3
. 4
. 5
. 6
. 7
. 8
. 9
. SkDH aa bb bb aa aa bb bb bb 9 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 aa cc cc BC cc BC UGPP bb 19 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 а.к. aa aa aa bb aa aa 90 190 а. ) 1 2,3 4 5,13 7 8,11 9 10 12 NF 1 0,0 0,0 1 1,? * 4 1 3
| . | Генотипы очаговых особей . | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Локус . | 1 . | 2 . | 3 . | 4 . | 5 . | 6 . | 7 . | 8 . | 9 . | |||||||||
| SkDH | aa | bb | bb | aa | aa | bb | bb | bb | 9 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 | aa | cc | cc | BC | cc | BC | |||
| UGPP | bb | 19 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 а.к. | aa | aa | aa | bb | aa | aa | 90 190 а. )1 | 2,3 | 4 | 5,13 | 7 | 8,11 | 9 | 10 | 12 | |
| NF | 1 | 0,0 | 0,0 | 1 | 1,? * | 4 | 1 | 3 | ||||||||||
Asplenium scolopendrium генотипов (очаговых особей), использованных в эксперименте, по данным анализа 11 аллозимных локусов и места их колонизации
| . | Генотипы очаговых особей . | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Локус . | 1 . | 2 . | 3 . | 4 . | 5 . | 6 . | 7 . | 8 . | 9 . | |||||||||
| SkDH | aa | bb | bb | aa | aa | bb | bb | bb | 9 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 | aa | cc | cc | BC | cc | BC | |||
| UGPP | bb | 19 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 а.к. | aa | aa | aa | bb | aa | aa | 90 190 а. )1 | 2,3 | 4 | 5,13 | 7 | 8,11 | 9 | 10 | 12 | |
| NF | 1 | 0,0 | 0,0 | 1 | 1,? * | 4 | 1 | 3 | ||||||||||
| . | Генотипы очаговых особей . | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Локус . | 1 . | 2 . | 3 . | 4 . | 5 . | 6 . | 7 . | 8 . | 9 . | |||||||||
| SkDH | aa | bb | bb | aa | aa | bb | bb | bb | 9 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 | aa | cc | cc | BC | cc | BC | |||
| UGPP | bb | 19 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 9019 а.к. | aa | aa | aa | bb | aa | aa | 90 190 а. )1 | 2,3 | 4 | 5,13 | 7 | 8,11 | 9 | 10 | 12 | |
| NF | 1 | 0,0 | 0,0 | 1 | 1,? * | 4 | 1 | 3 | ||||||||||
Кресты устанавливали в 6-сантиметровых чашках Петри на той же среде, что и ранее.Все экспериментальные единицы были перенесены в теплицу (Ботанический сад Утрехта) после пересадки (март 2008 г.). Помимо герметизации всех чашек Петри с помощью Parafilm ® , потеря влаги была дополнительно предотвращена путем установки относительной влажности воздуха в теплице на 90%. Три натриевые лампы мощностью 400 Вт были подвешены прибл. 2 м за время эксперимента, чтобы увеличить естественный фотопериод до 16 часов в день. Теплица отапливалась до 18 ° C днем и до 16 ° C ночью. Не было оборудования для охлаждения теплицы, поэтому можно было легко достичь более высоких температур, особенно летом.Ткань была подвешена вокруг установки, чтобы прямые солнечные лучи не повредили гаметофиты. Измерения света показали значительную неоднородность интенсивности света по всему участку эксперимента (данные не показаны), поэтому вся установка была рандомизирована каждые две недели. При наблюдении за первыми гаметангиями (через 12 недель после посева) все экспериментальные единицы поливали каждые две недели стерильной водой. К обоим гаметофитам (если применимо) применялась вода, чтобы соединить их одной водной пленкой, тем самым облегчая свободное перемещение сперматозоидов между ними.Каждые две недели все гаметофиты основных растений проверяли на продукцию спорофитов и регистрировали смертность гаметофитов. После трансплантации за экспериментом наблюдали еще 32 недели (т.е. до 38 недель после посева).
2″ data-legacy-id=»s3″> РЕЗУЛЬТАТЫ
11 проверенных ферментных систем выявили восемь полиморфных локусов в восьми отобранных европейских популяциях, в то время как пять из этих локусов были полиморфными в Kuinderbos.Восемь полиморфных локусов выявили в общей сложности 20 аллелей, пять из которых принадлежали отдельным популяциям. У населения Куиндербоса не было частных аллелей. Представители Kuinderbos вместе составляют ок. 95% всех нечастных аллелей (75% от общего числа) обнаружены в семи контрольных популяциях со всей Европы. Следовательно, большая часть генетической изменчивости вида была представлена в эксперименте. Особи, которые использовались в качестве основных растений в эксперименте, представляют девять различных генотипов (таблица 1).
Первые спорофиты были зарегистрированы через 14 недель после посева, и их количество постоянно росло до 22 недели (рис. 1А). К тому времени гибель гаметофитов существенно ограничила возможности для дальнейших оплодотворений (Приложение 1). Из-за отсутствия охлаждающих устройств в теплице максимальные дневные температуры воздуха регулярно превышали 30 ° C, начиная с 14 недели (данные не показаны). В конце эксперимента (38 неделя) почти 18% очаговых гаметофитов дали спорофит.
Рис.1.
Среднее производство спорофитов с течением времени для различных уровней инбридинга (A) для всех родительских спорофитов и для родительских спорофитов (B) на всех уровнях инбридинга. I — интрагаметофитное самоопыление; II, внутриплощадочные кроссы; III, межузельные кресты. Планки погрешностей представляют собой стандартную ошибку среднего ( n = 13).
Рис. 1.
Среднее производство спорофитов с течением времени для разных уровней инбридинга (A) для всех родительских спорофитов и для родительских спорофитов (B) на всех уровнях инбридинга.I — интрагаметофитное самоопыление; II, внутриплощадочные кроссы; III, межузельные кресты. Планки погрешностей представляют собой стандартную ошибку среднего ( n = 13).
В ходе эксперимента наблюдались многочисленные случаи абортов эмбрионов. Эмбрионы начинались как скопления клеток внутри архегонии, вызывая видимое набухание внутри гаметофита. Эмбрионы считались абортированными, если они не давали корней или листьев. К сожалению, систематически такие аборты не регистрировались.Полиэмбриония, то есть появление двух или более спорофитов из одного гаметофита, наблюдалась только при двух скрещиваниях, оба на гаметофитах, происходящих из очагового спорофита сайта KB-12.
Продукция спорофитов заметно различалась на трех уровнях инбридинга (Рис. 1A и Таблица 2). В целом, продукция спорофитов была самой высокой при межсайтовых скрещиваниях (III), при этом почти 30% очаговых гаметофитов в конечном итоге продуцировали спорофит. Скрещивания внутри участка (II) были менее чем вдвое менее успешными (прибл.14%), тогда как интрагаметофитное самоопыление (I) имело наименьший процент успеха. Тем не менее, в последнем варианте (I) средняя продукция спорофитов все еще составляла почти 10%. Гаметофиты только трех из 13 родительских спорофитов не продуцировали спорофитов изолированно (KB-8, -11 и -13). Все остальные продуцировали несколько спорофитов.
Таблица 2.Бинарный логистический регрессионный анализ продукции спорофитов через 16, 20, 24 и 28 недель после посева (для каждого анализа показаны критерии значимости Nagelkerke R 2 и Вальда для уровня инбридинга, родительских спорофитов и гаметофитов)
| . | . | Уровень инбридинга . | Родительский спорофит . | Смертность гаметофитов * . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Неделя . | р 2 . | Wald . | д.ф. . | Sig. . | Wald . | д.ф. . | Sig. . | Wald . | д.ф. . | Sig. . |
| 16 | 0 · 208 | 28 · 366 | 2 | <0 · 0005 | 24 · 099 | 12 | 0 · 0201| 191 | 1 | 0 · 999 | |
| 20 | 0 · 217 | 42 · 113 | 2 | <0 · 0005 | 39 · 591 | 12 | 0 < | <0 · 0005 | 1 | 0 · 999 |
| 24 | 0 · 225 | 41 · 597 | 2 | <0 · 0005 | 52 · 143 | <0 · 0005 | 6 · 090 | 1 | 0 · 014 | |
| 28 | 0 · 233 | 40 · 859 | 2 | <0 · 0005010 | 38612 | <0 · 0005 901 91 | 13,369 | 1 | <0,0005 | |
| . | . | Уровень инбридинга . | Родительский спорофит . | Смертность гаметофитов * . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Неделя . | р 2 . | Wald . | д.ф. . | Sig. . | Wald . | д.ф. . | Sig. . | Wald . | д.ф. . | Sig. . |
| 16 | 0 · 208 | 28 · 366 | 2 | <0 · 0005 | 24 · 099 | 12 | 0 · 0201| 191 | 1 | 0 · 999 | |
| 20 | 0 · 217 | 42 · 113 | 2 | <0 · 0005 | 39 · 591 | 12 | 0 < | <0 · 0005 | 1 | 0 · 999 |
| 24 | 0 · 225 | 41 · 597 | 2 | <0 · 0005 | 52 · 143 | <0 · 0005 | 6 · 090 | 1 | 0 · 014 | |
| 28 | 0 · 233 | 40 · 859 | 2 | <0 · 0005010 | 38612 | <0 · 0005 901 91 | 13,369 | 1 | <0,0005 | |
Бинарный логистический регрессионный анализ продукции спорофитов через 16, 20, 24 и 28 недель после посева (для каждого анализа показаны критерии значимости Nagelkerke R 2 и Вальда для уровня инбридинга, родительской смертности спорофитов и гаметофитов)
| . | . | Уровень инбридинга . | Родительский спорофит . | Смертность гаметофитов * . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Неделя . | р 2 . | Wald . | д.ф. . | Sig. . | Wald . | д.ф. . | Sig. . | Wald . | д.ф. . | Sig. . |
| 16 | 0 · 208 | 28 · 366 | 2 | <0 · 0005 | 24 · 099 | 12 | 0 · 0201| 191 | 1 | 0 · 999 | |
| 20 | 0 · 217 | 42 · 113 | 2 | <0 · 0005 | 39 · 591 | 12 | 0 < | <0 · 0005 | 1 | 0 · 999 |
| 24 | 0 · 225 | 41 · 597 | 2 | <0 · 0005 | 52 · 143 | <0 · 0005 | 6 · 090 | 1 | 0 · 014 | |
| 28 | 0 · 233 | 40 · 859 | 2 | <0 · 0005010 | 38612 | <0 · 0005 901 91 | 13,369 | 1 | <0,0005 | |
| . | . | Уровень инбридинга . | Родительский спорофит . | Смертность гаметофитов * . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Неделя . | р 2 . | Wald . | д.ф. . | Sig. . | Wald . | д.ф. . | Sig. . | Wald . | д.ф. . | Sig. . |
| 16 | 0 · 208 | 28 · 366 | 2 | <0 · 0005 | 24 · 099 | 12 | 0 · 0201| 191 | 1 | 0 · 999 | |
| 20 | 0 · 217 | 42 · 113 | 2 | <0 · 0005 | 39 · 591 | 12 | 0 < | <0 · 0005 | 1 | 0 · 999 |
| 24 | 0 · 225 | 41 · 597 | 2 | <0 · 0005 | 52 · 143 | <0 · 0005 | 6 · 090 | 1 | 0 · 014 | |
| 28 | 0 · 233 | 40 · 859 | 2 | <0 · 0005010 | 38612 | <0 · 0005 901 91 | 13,369 | 1 | <0,0005 | |
Заметные различия наблюдались и среди родительских спорофитов (рис.1Б и таблица 2). В межсайтовых скрещиваниях (III) различия в продукции спорофитов среди фокальных особей стали наиболее интересными, так как в этой обработке они были объединены с гаметофитами из случайно назначенных мест колонизации. Таким образом, даже несмотря на то, что гаметофиты каждого очагового спорофита были представлены одной и той же палитрой партнеров, все же возникали заметные различия в производительности (рис. 2). Кроме того, способность KB-4 к самоопылению (I) была в 3 раза выше, чем у любого другого спорофита, и конкурировала с собственными характеристиками при межсайтовых (III) скрещиваниях (рис.2).
Рис. 2.
Продукция спорофитов через 20 недель после посева для всех 13 родительских спорофитов и всех обработок. n = 20 для всех комбинаций, кроме отмеченных звездочкой, где n = 19.
Рис. 2.
Продукция спорофитов через 20 недель после посева для всех 13 родительских спорофитов и всех обработок. n = 20 для всех комбинаций, кроме отмеченных звездочкой, где n = 19.
Различия между уровнями инбридинга и родительских спорофитов, хотя и учитывали смертность гаметофитов, были очень значительными с течением времени (Таблица 2). Не было значительного взаимодействия между уровнем инбридинга и родительским спорофитом (не показано). Попарное сравнение уровней инбридинга показало значительную разницу между внутрисайтовыми (II) и межсайтовыми (III) скрещиваниями [например, неделя 20; соотношение шансов: 3 · 29; 95% ДИ: (1 · 96: 5 · 49), P <0,001].Различия между внутригаметофитным самоопылением (I) и внутрисайтовым скрещиванием (II) были значительными только после исключения родительского спорофита KB-4 [например, неделя 20; соотношение шансов: 2 · 74; 95% ДИ: (1 · 22: 6 · 15), . P = 0,030; в том числе КБ-4, Р = 0,168].
9″ data-legacy-id=»s4a»> Инбридинговая депрессия в условиях стресса
В настоящем эксперименте смертность гаметофитов сильно ограничивала оплодотворение через 22 недели.Однако, на наш взгляд, это естественная ситуация. Показано, что уровень смертности гаметофитов в полевых условиях очень высок, при этом большинство из них погибает в течение нескольких месяцев (Peck et al. , 1990). Точно так же предварительные результаты систематического изучения динамики популяции Polystichum aculeatum в Kuinderbos показывают, что большинство гаметофитов присутствует в течение <4 месяцев (G. A. de Groot, unpubl. Res.). По прошествии этого времени они либо сумели произвести спорофиты, либо исчезли без видимого репродуктивного успеха.Этот временной интервал короче, чем период между посевом и моментом, когда смертность гаметофитов стала ограничивающим фактором в эксперименте.
Низкая наблюдаемая в эксперименте продукция спорофитов, вероятно, является следствием высоких температур воздуха в теплице (дневной максимум> 30 ° C). Это привело к высокой смертности гаметофитов, достигшей пика на 24 неделе, что фактически препятствовало дальнейшему оплодотворению. Такие экстремальные температуры вряд ли будут регулярно наблюдаться в затененных местах на большей части Европы.Однако многие популяции A. scolopendrium в Нидерландах живут в стенах, и эта среда может быть более экстремальной в этом отношении. Кроме того, другие факторы, такие как летняя засуха (Cinquemani Kuehn and Leopold, 1992), могут сделать окружающую среду столь же стрессовой для гаметофитов A. scolopendrium . Более того, ожидается, что инбридинговая депрессия будет наиболее очевидной в стрессовых условиях (Армбрустер и Рид, 2005; Гудвилли и др. , 2005) из-за абиотического стресса (Дудаш, 1990) или межвидовой конкуренции (Плюсс и Стёклин, 2004).Настоящие наблюдения внутригаметофитного самоопыления при высоких показателях смертности гаметофитов, таким образом, усиливают его потенциальное значение в природе, и способность вида к воспроизводству таким образом в идеальных условиях может быть выше.
Развитие и половая экспрессия гаметофитов в лабораторных экспериментах могут отличаться от моделей роста в полевых условиях (Cousens, 1979; Pangua et al. , 1994; Ranker and Houston, 2002). Например, в стрессовых условиях гаметофиты имели тенденцию становиться самцами в Matteuccia struthiopteris , в то время как относительно мягкая среда давала больше самок [Prantl, 1881 (цитируется по Näf, 1979)].Однако природные условия также различаются, и, таким образом, ожидается, что модели выражения пола будут также различаться в разных популяциях на местах (см., Например, данные Ranker and Houston, 2002). Тем не менее, настоящий эксперимент показывает, что помимо ауткроссинга, у A. scolopendrium возможно внутригаметофитное самоопыление, и это интерпретируется как свидетельство смешанной системы спаривания.
7″ data-legacy-id=»s4c»> Внутривидовая изменчивость репродуктивного успеха В настоящем эксперименте были обнаружены большие различия в репродуктивной способности среди генотипов A. scolopendrium . Хотя системы спаривания были экспериментально изучены у различных видов папоротников (например, Korpelainen, 1996; Lott et al. , 2003; Pangua et al. , 2003; Flinn, 2006), в нескольких исследованиях сообщалось о внутривидовых вариациях (но см. Peck et al., 1990; Suter et al. , 2000). Большие различия в скорости самоопыления наблюдались среди популяций при анализе генетической изменчивости папоротников (например, Soltis and Soltis, 1987, 1992; Ranker, 1992; Ranker et al. , 2000; Pryor et al. , 2001), как было также обнаружено у покрытосеменных (например, Arabis alpina ; см. Ansell et al. , 2008) и мохообразных (например, Leucodon sciuroides ; см. Cronberg, 2000).
В настоящем исследовании показатель успешности внутригаметофитного самоопыления у A.scolopendrium варьировала от нуля до 45%. По сравнению с исследованием Peck et al. (1990), этот диапазон охватывает вариации внутриигаметофитной скорости самоопыления у восьми из десяти изученных ими видов. Поскольку условия окружающей среды в разных экспериментах различаются, результаты трудно сравнивать напрямую между исследованиями. Тем не менее, это указывает на то, что исследования системы разведения должны включать как можно более широкий выбор внутривидовых вариаций, чтобы иметь возможность делать выводы на уровне видов.Кроме того, настоящие результаты ставят под сомнение слишком простую дихотомическую классификацию видов папоротников как преимущественно самоопыляющихся или ауткроссинговых. Результаты требуют всестороннего экспериментального исследования с участием нескольких видов и генотипов папоротников внутри видов, чтобы оценить различия в производительности внутри видов по сравнению с различиями между видами.
2″> БЛАГОДАРНОСТИ Мы благодарим Фреда Сислинга (Ботанический сад Утрехта) за его помощь во время эксперимента и Пола Вестерса (Университетский медицинский центр Утрехта) за статистические советы. Работа поддержана Фондом Микеля (Утрехтский университет).
6″> ПРИЛОЖЕНИЕ Смертность гаметофитов в эксперименте с течением времени, усредненная по уровню инбридинга. I — интрагаметофитное самоопыление; II, внутриплощадочные кроссы; III, межузельные кресты.
© Автор, 2010. Опубликовано издательством Oxford University Press от имени компании Annals of Botany. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
. Жизненный цикл папоротника: размножение растений без цветов и семян — класс CLEP [видео 2021]
Обзор: чередование поколений
Это чередование поколений представляет собой жизненный цикл, который включает как диплоидные, так и гаплоидные многоклеточные стадии.Помните, что диплоид означает два набора хромосом и обычно обозначается сокращенно как «2N», потому что N обозначает хромосомы. В диплоидных клетках по одной копии хромосом от каждого родителя. Например, у людей вы получаете одну копию хромосом от отца и одну копию хромосом от мамы.
Та же идея встречается и в растениях. Каждая диплоидная клетка содержит одну копию хромосом от мужского родителя и одну копию хромосом от женского родителя. Гаплоид означает один набор хромосом и обычно обозначается аббревиатурой «N», потому что существует только одна копия хромосом.
Диаграмма смены поколений
Рассмотрим схему основной идеи смены поколений. На этой диаграмме видно, что жизненный цикл разбит на N вверху и 2N внизу. Помните, что гаметофит содержит гаплоидные клетки, а спорофит — диплоидные клетки. Ранее мы использовали garblinx, чтобы проиллюстрировать странность этого. Помните, что диплоидный, или 2N, организм выглядит так.
Диаграмма, показывающая диплоидный организм
Однако, когда он переходит в гаплоидную стадию, гарблинкс выглядит совершенно иначе! Два из этих гаплоидных организмов собираются вместе и спариваются, чтобы произвести новый организм 2N, похожий на первый гарблинкс. Затем этот гарблинкс в конечном итоге произведет новый гаплоидный организм и так далее.
Теперь, когда мы рассмотрели основные аспекты этого жизненного цикла, давайте посмотрим, как папоротники проходят этот цикл.Мы начнем с гаплоидной стадии, как делали раньше, а затем перейдем к стадии доминантного диплоида.
Гаплоидная стадия
Мы будем использовать эту диаграмму, чтобы проиллюстрировать жизненный цикл папоротников.
Схема жизненного цикла папоротника
Папоротник начинается с гаплоидной стадии в виде спор. Спора подвергнется митозу, чтобы создать множество идентичных гаплоидных клеток. Эта группа клеток называется гаметофитом .На нашей диаграмме видно, что спора перерастает в гаметофит. Большинство гаметофитов папоротника выглядят как листья в форме сердца и меньше вашего мизинца.
Интересно, что этот гаметофит, несмотря на невысокие размеры и структуру, способен к фотосинтезу. Это означает, что гаметофит может вырабатывать собственную энергию и расти независимо.
После развития и роста в качестве гаметофита спора готова производить гамет . Гаметы образуются в определенной области гаметофита. Gametangia — это органы гаметофитов, производящие гаметы.
Есть два типа гаметангий: один производит яйцеклетки, а другой — сперматозоиды. архегонии — это женские гаметангии, которые производят одно яйцо за раз. Антеридии — мужские гаметангии, которые производят много сперматозоидов. Вы можете вспомнить, какие гаметангии мужские, а какие женские, потому что пыльники — это мужские структуры в цветках, производящие сперму. Если вы знаете, что пыльники производят сперму, вы можете помнить, что антеридии также производят сперму и, следовательно, являются мужскими.
На нашей диаграмме показаны мужские и женские структуры одного и того же гаметофита. Это означает, что гаметофит папоротника бисексуален, потому что он содержит как мужские, так и женские структуры. Антеридии расположены ближе к верхушке сердцевидного листа. Архегонии расположены в верхней части гаметофита. С генетической точки зрения, для гамет лучше всего смешиваться между разными гаметофитами, чтобы начать диплоидную стадию. Если гаметы происходят от одного и того же растения, генетического разнообразия не будет.Однако, если яйцеклетка и сперма происходят от разных растений, это увеличит генетическую изменчивость, что, скорее всего, повысит шансы растения на выживание.
Диплоидная стадия
Диплоидная — и доминирующая — стадия жизни начнется после того, как произойдет оплодотворение. Помните, что оплодотворение — это слияние яйцеклетки и спермы. И яйцеклетка, и сперматозоид гаплоидны, поэтому, когда они сливаются вместе, мы получаем диплоидную клетку. Эта новая клетка будет продолжать митоз и расти. В конце концов, он станет зрелым спорофитом, похожим на известные вам папоротники.У зрелого спорофита много разных структур, но мы остановимся на тех, которые участвуют в размножении.
На нижней стороне листьев папоротника вы можете найти красные или коричневые точки. Эти точки называются сори и представляют собой группы спорангиев.
Что такое спорангии? Подобно тому, как у гаметофитов есть гаметангии, производящие гаметы, у спорофитов есть спорангии, производящие споры. Спорангии — это органы, производящие споры в процессе мейоза.
Фото сори на нижней стороне листьев папоротника
Внутри каждого спорангия есть клетки, которые фактически делятся и становятся спорами. Эти спороцитов представляют собой диплоидные клетки, которые подвергаются мейозу с образованием гаплоидных спор. Эти споры микроскопические, то есть очень маленькие. Хотя вы можете увидеть группы спорангиев на нижней стороне листьев папоротника, вы не можете увидеть споры без помощи микроскопа.После высвобождения эти споры начнут гаплоидную стадию жизненного цикла.
Резюме урока
Папоротники — уникальные растения, потому что они не дают семян. Эти сосудистые растения адаптировались к жизненному циклу, в котором используются споры, а не семена. Это чередование поколений у папоротников означает, что есть многоклеточные стадии, которые бывают гаплоидными и диплоидными.
Сначала мы рассмотрели гаплоидную стадию, которая начинается как спора и развивается в гаметофит .Гаметофит папоротника выглядит как небольшой лист в форме сердца и является обоеполым, он содержит как мужские, так и женские органы, производящие гамет. гаметангии , производящие сперму, называются антеридиями , а те, которые производят яйцеклетки, называются архегониями .
Как только яйцеклетка оплодотворяется спермой, папоротник переходит в диплоидную стадию жизненного цикла. Эта новая диплоидная клетка развивается в зрелый спорофит, который представляет собой структуру, о которой вы думаете, когда думаете о папоротниках.У спорофита есть уникальные структуры, называемые спорангии , расположенные на нижней стороне некоторых листьев. Эти структуры содержат спороцита , которые продуцируют споры . Эти споры снова начинают гаплоидную стадию.
Результат обучения
В конце этого урока вы сможете описать этапы уникального жизненного цикла папоротников.
Яванский папоротник: полное руководство по уходу (виды, посадка и размножение)
Яванский папоротник, с научной точки зрения известный как Microsorum pteropus , является классическим и очень популярным аквариумным растением.
Этот нежный на вид папоротник придает аквариуму красивый вид. Он широко используется из-за медленного роста, уникальной формы и простоты воспроизводства и ухода.
Может содержаться с самыми разными рыбами и не требует строгих параметров воды, поэтому так нравится многим людям. Это одно из самых простых растений для начинающих.
В этой статье мы расскажем о происхождении и внешнем виде растения, о том, как его сажать и поддерживать, о методах размножения и подходящих помощниках по аквариуму.Прежде чем продолжить чтение, вы можете взглянуть на это краткое резюме.
Категория Рейтинг Семья: Polypodiaceae Уровень ухода: Easy Скорость роста до 13,5 дюймов Минимальный размер бака: 10 галлонов Условия воды: 68-82 o F, pH 6.0-7,5, KH 3-8 Освещение: От слабого до умеренного Размножение: Придаточное растение / корневище Размещение: Середина к фону Яванский папоротник — член семейства полиподиевых, насчитывающего более 60 различных родов. Microsorum — это род этого семейства, которое включает более 50 видов тропических папоротников.
Пожалуй, одним из самых популярных видов рода Microsorum является Microsorum pteropus, также известный как Яванский папоротник.
Произошло из Юго-Восточной Азии и представляет собой растение в джунглях, которое обычно растет на камнях, земле и вокруг грузовиков с деревьями вдоль ватерлинии паров и водопадов. Он также растет в тропических дождевых лесах, как трава. Он может расти как при полностью погруженном, так и частично погруженном в воду.
Ява Папоротник давно является хобби, и с годами были выведены новые разновидности.Самые популярные из них, которые вы найдете в рыбных магазинах, — это узколистный, игольчатый, трезубец и винделов.
Это относительно дешево; Вы можете купить небольшое растение примерно за 4-5 долларов. Иногда они уже прикреплены к корягу, что позволяет легко попасть прямо в аквариум. Растения, которые уже растут на древесине, как правило, стоят дороже, в зависимости от размера, вы можете рассчитывать заплатить более 20 долларов.
Когда вы покупаете их, они обычно будут около 3-5 дюймов в длину. Убедитесь, что вы выбрали здоровое растение без коричневой окантовки и зеленых, здоровых на вид листьев.
Не покупайте пачку, иногда магазины просто собирают вместе партию черенков, и у них отсутствуют корневища.
Это отличное растение, если у вас ограниченный бюджет. Практически невозможно убить это растение, если вы не предпринимаете активных действий!
Внешний вид папоротника Явы
Папоротник Явы — традиционное зеленое аквариумное растение, состоящее из двух основных компонентов; корневище и листья.
Корневища действуют как якорь и представляют собой темно-коричневые, похожие на волоски струны, которые прикрепляются ко многим различным поверхностям.
Листья очень выносливые и имеют кожистую текстуру различных уникальных форм, от кустистых до колючих. Он бывает зеленого цвета от среднего до темно-зеленого. Обычно чем выше освещение, тем темнее зеленый цвет. На некоторых зрелых листьях появляется несколько черных / коричневых крошечных круглых бугорков (один из способов размножения, который мы вскоре обсудим), а иногда и несколько черных прожилок, проходящих через листья.
Растение может достигать высоты около 13,5 дюймов и может достигать 6-8 дюймов в ширину.Это делает его идеальным для самых разных аквариумов, от больших общественных аквариумов до густо засаженных или даже демонстрационных.
У оригинального растения есть варианты, которые влияют на размер и форму листьев. Мы рассмотрим некоторые из самых популярных разновидностей аквариумов; однако есть и другие, менее известные разновидности.
- Узколистный Яванский папоротник: У этого растения узкие листья, которые растут под более крутым углом, чем у обычного Microsorum pteropus .Листья вырастают примерно до 4-8 дюймов, а растение может вырасти до 12 дюймов.
- Игольчатый папоротник Ява: Это растение имеет еще более тонкие листья и меньше по размеру, чем узколистное растение. Он может вырасти до 6 дюймов в высоту. Настоящий игольчатый лист Ява довольно редко встречается в торговле.
- Трезубец Ява Папоротник: Это один из наиболее необычных вариантов, у него перистые лопастные листья с 2-5 долями с каждой стороны. Он короче узкого листа, но растет быстрее и имеет больше вилок на каждом листе.
- Windelov Ява Папоротник: Уникальный вариант с мелко разветвленными кончиками листьев. Обычно он вырастает примерно до 8 дюймов в высоту.
Java Fern Care
Microsorum pteropus Требования к аквариуму
Как и рыбы, водные растения процветают, когда условия аквариума максимально соответствуют их естественным условиям.
В своем естественном климате папоротник Явы растет вдоль ручьев и часто встречается у движущихся водоемов. Фильтры и насадки обеспечивают достаточное количество кислорода для этих растений в аквариуме — им не нужно много дополнительного CO2.
Их также можно найти на дне джунглей, где мягкая кислая вода брызгает на листья из ручьев и водопадов. Для воспроизведения этих условий рекомендуется значение pH от 6,0 до 7,0 с жесткостью 3-8dGH.
Это чрезвычайно выносливое растение, которому не нужны особые условия для роста. Ему не требуется яркое освещение, хороший субстрат или большое количество питательных веществ, поэтому это растение — мечта.
Чтобы он рос, вам не нужен красивый аквариум; он будет расти при самых простых настройках.По размеру его можно выращивать в резервуарах размером до 10 галлонов. Мы не рекомендуем помещать его в емкости меньшего размера. Он будет расти в большинстве условий освещения; наиболее предпочтительны приглушенные люминесцентные лампы или лампы накаливания. Не делайте освещение слишком сильным — они станут коричневыми и прозрачными, если освещение будет слишком сильным.
В этом случае уменьшите освещение или выключите его на несколько дней, пока растение не восстановится. В дикой природе эти растения фактически растут в тени и не подвергаются воздействию яркого света.
В идеале, при использовании ламп 5000-7000K на каждый галлон в вашем резервуаре требуется 1,5 / 2 Вт света.
С точки зрения температуры, растение будет расти в холодной воде и в тропических резервуарах, его предпочтительная температура составляет 68-82 o F.
Он также идеально подходит для резервуаров с голым дном, поскольку не требует никакого субстрата. Если вы все же хотите добавить субстрат в аквариум с растением, выберите все, что вам нравится; это растение не извлекает питательные вещества из субстрата.
Интересно, что некоторые люди настаивают на том, что это растение может выжить в солоноватой воде, и пока оно будет жить какое-то время, оно, скорее всего, не будет расти и в конечном итоге умрет.
Как сажать Явский папоротник
Явский папоротник, привязанный к дереву Явский папоротник имеет корневища, которые не любят закапывать. Растение будет расти очень медленно или может вообще не расти, если вы его закопаете.
Вместо того, чтобы закапывать его, вам нужно прикрепить его к корягу, камням или чему-то подобному.
Шероховатые поверхности лучше гладкие. Все, что угодно, например, лавовый камень или коряги, подойдет идеально. Если поверхность объекта слишком гладкая (например, галька или стекло), потребуется больше времени для прикрепления, но обычно он все равно в конечном итоге прикрепляется.Желательно использовать большой кусок коряги, чтобы у него было достаточно места для разложения, но можно использовать и маленькие кусочки.
Чтобы обезопасить растение, привяжите корни с помощью рыболовной проволоки к выбранному вами объекту. Черный цвет предпочтительнее использовать, так как он соответствует цвету корней. Некоторые люди предпочитают использовать резинки или стяжки, однако они обычно видны снаружи вашего бака.
Вы обнаружите, что через несколько недель корни прикрепятся к поверхности того, к чему вы их прикрепили.
Как только корни закреплены, вы можете удалить их стяжки; вы можете сделать это, если выбрали хорошо заметный галстук. Вам не нужно снимать леску и т. Д., Если вы не хотите, так как растение будет прикрывать ее по мере роста.
Яванский папоротник может вырасти довольно большими и с особенно широкими листьями; Поэтому рекомендуется сажать его посередине или ближе к задней части резервуара, если вы не пытаетесь добиться эффекта джунглей.
Если вы поместите его в передней части резервуара, он скроет все остальное, что у вас там есть.
Вы можете сажать его отдельно, группами или другими растениями.
Вместо этого вы можете захотеть поплавать это растение, однако его корневища будут расти все дольше и дольше, пока оно не найдет, к чему прикрепиться.
Уход и уход
Папоротник Ява — одно из самых простых в уходе растений; Если у вас когда-либо возникали проблемы с выращиванием аквариумных растений, попробуйте это.
Вы можете обнаружить, что вашему растению нужно время, чтобы начать расти, возможно, оно просто приспосабливается к условиям вашего аквариума и закладывает корневища.Как только это произойдет, он начнет расти.
То, как вы ухаживаете за своим, зависит от того, какой внешний вид вы хотите достичь в своем аквариуме. Если вы хотите, чтобы ваши растения были маленькими и отдельными, вам нужно будет удалить маленькие новые ростки, которые развиваются на листьях по мере их появления (мы расскажем, как это сделать в разделе о размножении).
Если вы хотите создать действительно густой рост, просто оставьте растение, чтобы оно продолжало расти. По мере развития новых проростков вместо того, чтобы удалять их для создания новых растений, оставьте их расти, чтобы добиться эффекта большой густоты.
Эти растения получают питательные вещества из воды через листья, поэтому удобрения не требуются. Однако, если вы хотите использовать удобрения для ускорения роста, вы можете это сделать. Не имея настоящих корней, они получают большую часть питательных веществ через листья из водной толщи, поэтому удобрения для субстрата будут бесполезны. Вместо этого добавляйте жидкое удобрение каждый раз при подмене воды.
Если на вашем растении появились ожоги, вы можете обрезать растение, чтобы удалить их.
Из-за медленного роста вы можете обнаружить, что вам нужно делать только одну большую обрезку в год.Обрежьте листья как можно ближе к корневищу.
Прелесть Java Fern, прикрепляющегося к камням и корягам, заключается в том, что когда дело доходит до очистки аквариума, вы можете перемещать его во время очистки. Это особенно привлекательно для аквариумов, содержащих грязную рыбу.
N.B. Когда вы чистите свой аквариум или проводите какое-либо другое обслуживание, это растение необходимо постоянно поддерживать во влажном состоянии, поэтому либо держите его под водой, либо используйте распылитель, чтобы листья оставались влажными.
Общие проблемы
Многие новички думают, что все растения нужно сажать в субстрат.Это распространенная ошибка, которая приведет к смерти Java Fern. Вы обнаружите, что он выживет в течение нескольких недель в субстрате, но затем он начнет становиться коричневым, что является признаком того, что он умирает.
Вторая проблема, с которой сталкиваются люди, заключается в том, что они обнаруживают, что растение не растет. Отсутствие роста в первые несколько недель — это нормально. Если он все еще не растет, вы можете рассмотреть возможность использования удобрений, как указано выше.
Другая распространенная ошибка, которую люди совершают с этим растением, — это принимать черные / коричневые пятна на листьях за нездоровое растение.Эти черные круги — то место, откуда будут расти новые ростки, так что не волнуйтесь! Черные пятна, которые не развиваются в новые растения, могут быть признаком дефицита азота; другие пятна могут указывать на ожоги, поэтому выключите свет.
Последняя проблема, которую мы рассмотрим, — это плавление Java Fern. Для этого характерны большие коричневые пятна, которые, в свою очередь, вызывают гниение и размягчение растения.
Обычно это происходит из-за того, что растениям не хватает питательных веществ, слишком много света или слишком много сине-зеленых водорослей в вашем аквариуме.
В качестве примечания: если у вас есть немного водорослей, вы можете подумать о том, чтобы нанять бригаду по уборке, которая поможет держать их в курсе.
Ничто из этого не должно быть проблемой, если вы правильно настроили резервуар, правильно следуете приведенному выше разделу по посадке и уходу и регулярно меняете воду.
Размножение папоротника на Яве
Фото ictheostega. Размножение Java Fern очень просто и не требует каких-либо особых условий или работы с вашей стороны.
Есть несколько способов размножения:
- Во-первых, вы можете разрезать корневище пополам и заново засеять срезанные секции.Затем отдельные растения продолжат расти.
- В качестве альтернативы можно подождать, пока на листьях растения разовьются крошечные папоротники Явы. Иногда они начинаются с черных пятен на нижней стороне листа.
Примерно через 2-3 недели из черных шишек начинают прорастать крошечные листья.
Их можно легко срезать острыми стерилизованными ножницами и переместить в другое место на растении, чтобы дать начало новому росту. По мере развития и созревания ростки снова начинают процесс роста.
Помните, что новый завод все равно нужно будет привязать так же, как мы обсуждали ранее.
Соседи и совместимость с папоротником Java в аквариуме
Рыба-ангел в аквариуме с растениями Это растение очень совместимо с огромным разнообразием рыб.
Даже травоядные рыбы, обычно питающиеся аквариумными растениями, стараются держаться от них подальше. Он также имеет очень жесткую структуру листьев, что делает его непривлекательным для рыбы.
Он совместим как с игривыми мелкими рыбками, так и с крупными, такими как цихлиды, тигровые оскары, сомы, арованы и другие крупные рыбы, которые обычно повреждают более чувствительные растения.
Однако делайте это с осторожностью в начале жизненного цикла растений. Его корни еще не так прочны; следовательно, он будет подвержен повреждениям и опрокидыванию, если вы решите разместить в нем крупную и более агрессивную рыбу.
Если вы все-таки живете с более крупной рыбой, прикрепите растение к корягу или камням, чтобы не повредить корни.
Яванский папоротник — очень выносливое, сильное и крепкое растение, которое трудно разорвать на части. Если вы хотите, чтобы растение выглядело гладким и красивым, вам не следует размещать его вместе с игривыми или мелкими рыбками.
Подходит ли Яванский папоротник для вашего аквариума (резюме)?
Если у вас пресноводный аквариум, то почти 100% вероятность, что это растение подойдет ему.
Если вы новичок в этом хобби или опытный ветеран, это растение обладает множеством замечательных качеств, которые делают его желанным аквариумным растением.
За ним очень легко ухаживать и поддерживать. Его легко размножать, и с ним можно содержать широкий спектр рыб.
Доступно несколько различных вариантов; тип, который вы выберете, будет зависеть от наличия и личных предпочтений.
Есть ли у вас опыт работы с Java Fern? Мы хотели бы услышать — напишите нам сообщение в разделе комментариев ниже…
Уникальная система соматического эмбриогенеза в древовидном папоротнике Cyathea delgadii Sternb: важность типа эксплантата и физических и химических факторов
Akula A, Becker D, Bateson M (2000) Высокоурожайный повторяющийся соматический эмбриогенез и восстановление растений в выбранном чайном клоне TRI-2025 путем временного погружения. Plant Cell Rep 19: 1140–1145
CAS
Статья
Google Scholar
Ambrósio ST, de Melo NF (2004) Взаимодействие между сахарозой и pH во время культивирования in vitro Nephrolepis biserrata (Sw.) Шотт (Pteridophyta). Acta Bot Bras 18: 809–813
Артикул
Google Scholar
Ambrožič-Dolinšek J, Camloh M (1997) Гаметофитная и спорофитная регенерация из почковой чешуи папоротника Platycerium bifurcatum (Cav.) C. Chr. in vitro. Ann Bot 80: 23–28
Статья
Google Scholar
Bharati SK, Manabendra DC, Behari MP (2013) Размножение in vitro у Pteridophytes: обзор.Int J Res Ayurveda Pharm 4: 297–303
Статья
Google Scholar
Bui LT, Hurst A, Irish EE, Cheng CL (2012) Влияние сахаров и этилена на апоспорию и регенерацию у Ceratopteris richardii . Am J Plant Sci 3: 953–961
CAS
Статья
Google Scholar
Chen J-T, Chang W-C (2002) Влияние условий культивирования тканей и характеристик эксплантов на прямой соматический эмбриогенез у Oncidium «Gower Ramsey».Культ растительных клеток, тканей и органов 69: 41–44
CAS
Статья
Google Scholar
Cordle AR, Irish EE, Cheng C-L (2007) Индукция апогамии у Ceratopteris richardii . Int J Plant Sci 168: 361–369
Статья
Google Scholar
Cordle AR, Bui LT, Irish EE, Cheng CL (2011) Апогамия и апоспория, вызванные лабораторными методами, у Ceratopteris richardii .В: Фернандес Х., Кумар А., Ревилла М.А. (ред.) Работа с папоротниками: проблемы и приложения. Springer Science + Business Media, LLC, Берлин, стр. 25–36
Google Scholar
Cunha A, Fernandes-Ferreira M (1999) Влияние параметров среды на соматический эмбриогенез из эксплантатов гипокотилей льна ( Linum usitatissimum L.). J Plant Physiol 155: 591–597
CAS
Статья
Google Scholar
Dykeman BW, Cumming BG (1985) Размножение страусиного папоротника in vitro ( Matteuccia struthiopteris ).Can J Plant Sci 65: 1025–1032
CAS
Статья
Google Scholar
Fernández H, Revilla MA (2003) In vitro Культивирование декоративных папоротников. Культ растительных клеток, тканей и органов 73: 1–13
Статья
Google Scholar
Фернандес Х., Бертран А.М., Санчес-Тамес Р. (1996) Влияние условий культивирования тканей на апогамию у Driopteris affinis sp. affinis . Культ растительных клеток, тканей и органов 45: 93–97
Статья
Google Scholar
Фернандес Х., Бертран А.М., Санчес-Тамес Р. (1999) Биологические и пищевые аспекты, связанные с размножением папоротников. Культ растительных клеток, тканей и органов 56: 211–214
Статья
Google Scholar
Ferreira DAT, Sattler MC, Carvalho CR, Clarindo WR (2015) Эмбриогенный потенциал незрелых зиготических зародышей Passiflora : новое достижение для размножения in vitro без регуляторов роста растений.Культ органа растительной клетки, ткани. DOI: 10.1007 / s11240-015-0796-1
Google Scholar
Филонова Л.Х., Божков П.В., фон Арнольд С. (2000) Путь развития соматического эмбриогенеза у Picea abies по данным покадровой съемки. J Exp Bot 51: 249–264
CAS
Статья
PubMed
Google Scholar
Gaj M (2004) Факторы, влияющие на индукцию соматического эмбриогенеза и регенерацию растений с особым упором на Arabidopsis thaliana (L.) Heynh. Регламент роста растений 43: 27–47
CAS
Статья
Google Scholar
Gow W-P, Chen J-T, Chang W-C (2010) Усиление прямого соматического эмбриогенеза и роста ростков из листовых эксплантатов Phalaenopsis путем регулирования периода культивирования и длины эксплантата. Acta Physiol Plant 32: 621–627
Статья
Google Scholar
Gray DJ, McColley DW, Compton ME (1993) Высокочастотный соматический эмбриогенез из покоящихся семядолей семян сортов Cucumis melo .J Am Soc Hort Sci 118: 425–432
Google Scholar
Hirsch AM (1975) Влияние сахарозы на дифференциацию вырезанной ткани листа папоротника на гаметофиты или спорофиты. Физиология растений 56: 390–393
PubMed Central
CAS
Статья
PubMed
Google Scholar
Hirsch AM (1976) Развитие апоспоровых гаметофитов и регенерированных спорофитов из эпидермальных клеток иссеченных листьев папоротника: анатомическое исследование.Am J Bot 63: 263–271
Статья
Google Scholar
Хименес В.М. (2005) Участие растительных гормонов и регуляторов роста растений в соматическом эмбриогенезе in vitro. Регламент роста растений 47: 91–110
Статья
Google Scholar
Камада Х., Кобаяши К., Кийосуэ Т., Харада Х. (1989) Стресс-индуцированный соматический эмбриогенез в моркови и его применение для производства синтетических семян.In vitro Cell Dev Biol 25: 1163–1166
Статья
Google Scholar
Камада Х, Исикава К., Сага Х, Харада Х (1993) Индукция соматического эмбриогенеза в моркови под действием осмотического стресса. Plant Tissue Cult Lett 10: 38–44
CAS
Статья
Google Scholar
Karami O, Deljou A, Esna-Ashari M, Ostad-Ahmadi P (2006) Влияние концентраций сахарозы на соматический эмбриогенез у гвоздики ( Dianthus caryophyllus L.). Sci Hortic 110: 340–344
CAS
Статья
Google Scholar
Курияма А., Ходжох Т., Сугавара Ю., Мацусима Х., Такеучи М. (1989) Метод быстрого роста в культуре гаметофитов Equisetum arvense с антеридиями. Физиология растительных клеток 30: 1189–1192
Google Scholar
Курияма А., Кобаяси Т., Маеда М. (2004) Получение спорофитных растений Cyathea lepifera , древовидного папоротника, из гаметофита, культивируемого in vitro.J Jpn Soc Hortic Sci 73: 140–142
Статья
Google Scholar
Liao YK, Wu YH (2011) In vitro размножение Platycerium bifurcatum (Cav.) C. Chr. через инициирование зеленого шаровидного тела. Bot Stud 52: 455–463
CAS
Google Scholar
Лу Х., Како С. (1995) Роль высоких концентраций сахара в индукции соматического эмбриогенеза семядолей огурца.Sci Hortic 64: 11–20
CAS
Статья
Google Scholar
Mahdavi-Darvari F, Mohd Noor N, Ismanizan I (2015) Эпигенетическая регуляция и генные маркеры как сигналы раннего соматического эмбриогенеза. Культ растительных клеток, тканей и органов 120: 407–422
CAS
Статья
Google Scholar
Martin KP, Sini S, Zhang C-L, Slater A, Madhusoodanan PV (2006) Эффективная индукция апоспории и апогамии in vitro в серебряном папоротнике ( Pityrogramma calomelanos L.). Plant Cell Rep 25: 1300–1307
CAS
Статья
PubMed
Google Scholar
Materi DM, Cumming BG (1991) Влияние углеводной депривации на омоложение, апоспорию и регенерацию у спорофитов страусиного папоротника ( Matteuccia struthiopteris ). Can J Bot 69: 1241–1245
Статья
Google Scholar
Menéndez V, Villacorta NF, Revilla MA, Gotor V, Bernard P, Fernández H (2006) Экзогенные и эндогенные регуляторы роста при апогамии у Dryopteris affinis (Lowe) Fraser-Jenkins sp. affinis . Plant Cell Rep 25: 85–91
Статья
PubMed
Google Scholar
Mikuła A, Pożoga M, Tomiczak K, Rybczyński J (2015) Соматический эмбриогенез в папоротниках: новая экспериментальная система. Plant Cell Rep 34: 783–794
PubMed Central
Статья
PubMed
Google Scholar
Moura IR, Simões-Costa MC, Garcia J, Silva MJ, Duarte MC (2012) Культивирование in vitro спор древесных папоротников из семейств Cyatheaceae и Dicksoniaceae .Acta Hortic 937: 455–461
Статья
Google Scholar
Murashige T, Skoog F (1962) Пересмотренная среда для быстрого роста и биоанализов с культурами тканей табака. Physiol Plant 15: 473–497
CAS
Статья
Google Scholar
Nakagawa H, Saijyo T, Yamauchi N, Shigyo M, Kako S, Ito A (2001) Влияние сахаров и абсцизовой кислоты на соматический эмбриогенез из мело ( Cucumis melo L.) расширенная семядоля. Sci Hortic 90: 85–92
CAS
Статья
Google Scholar
Nhut DT, Le BV, Fukai S, Tanaka M, Van KTT (2001) Влияние активированного угля, размера эксплантата, положения экспланта и концентрации сахарозы на регенерацию растений и побегов Lilium longiflorum через культивирование молодых стеблей. Регламент роста растений 33: 59–65
Статья
Google Scholar
Parajuli J, Joshi SD (2014) In vitro исследование эффектов гормонов роста на развитие спорофитов Cyathea spinulosa .Int J Biodivers Conserv 6: 247–255
Статья
Google Scholar
Pinto G, Park Y-S, Silva S, Neves L, Araújo C, Santos C (2008) Факторы, влияющие на поддержание, пролиферацию и прорастание вторичных соматических зародышей Eucalyptus globulus Labill. Культ организации тканей клеток растений 95: 69–78
CAS
Статья
Google Scholar
Raemakers CJJM, Jacobsen E, Visser RGF (1995) Вторичный соматический эмбриогенез и применение в селекции растений.Euphytica 81: 93–107
Статья
Google Scholar
Рагхаван В. (1989) Биология развития гаметофитов папоротников. Cambridge University Press, Cambridge, pp. 259–295
Книга
Google Scholar
Рашид А. (2009) Введение в мохообразные: разнообразие, развитие и дифференциация. В: Глава 11: смена поколений. VIKAS Publishing House PVT LTD, Нью-Дели, стр. 237–249
Рыбчинский Ю., Микула А. (2011) Биотехнология древовидных папоротников: от спор до спорофитов.В: Фернандес Х., Кумар А., Ревилла М.А. (ред.) Работа с папоротниками: проблемы и приложения. Springer Science + Business Media, Нью-Йорк, Дордрехт, Гейдельберг, Лондон, стр 135–147
Google Scholar
Смит Д.Л., Крикориан А.Д. (1990) Производство соматических проэмбрионов из иссеченных, раненых зародышей зиготической моркови на среде без гормонов: оценка влияния pH, этилена и активированного угля. Plant Cell Rep 9: 34–37
CAS
Статья
Google Scholar
Somer A, Arbesú R, Menéndez V, Revilla MA, Fernández H (2010) Исследования индукции спорофитов в папоротниках in vitro.Euphytica 171: 203–210
Статья
Google Scholar
Такамура Т., Танака М. (1996) Соматический эмбриогенез из этиолированного черешка цикламена ( Cyclamen persicum Mill.). Plant Tissue Lett 13: 43–48
CAS
Статья
Google Scholar
Tavares JE, Sussex IM (1968) Расширенный рост изолированных листовых пластинок Todea barbara .Planta 80: 113–128
CAS
Статья
Google Scholar
White RA (1971) Экспериментальные и эволюционные исследования спорофита папоротника. Bot Rev 37: 509–540
Статья
Google Scholar
Whittier DP, Pratt LH (1971) Влияние качества света на индукцию апогамии в отростках Pteridium aquilinum . Planta 99: 174–178
CAS
Статья
PubMed
Google Scholar
Whittier DP, Steeves TA (1960) Вызвание апогамии у папоротника папоротника.Can J Bot 38: 925–930
CAS
Статья
Google Scholar
Ян X, Чжан X (2010) Регулирование соматического эмбриогенеза у высших растений. Crit Rev Plant Sci 29: 36–57
CAS
Статья
Google Scholar
Ян-Лин С., Сун-Кван Х. (2012) Последние достижения в области эмбриогенеза однодольных и двудольных растений in vitro. В: Кен-Ичи Сато (ред) Эмбриогенез.ISBN: 978-953-51-0466-7, InTech. http://www.intechopen.com/books/embryogenesis/recent-advances-of-invitro-embryogenesis-of-monocotyledon-and-dicotyledon
Заваттьери М.А., Фредерико А.М., Лима М., Сабино Р., Арнхольдт-Шмитт Б. (2010) Индукция соматического эмбриогенеза как пример стресс-связанных реакций растений. Electron J Biotech 13. http://www.ejbiotechnology.cl/content/vol13/issue1/full/4/
Страница не найдена — Ботанический сад Калифорнийского университета
Сад открыт только по предварительному заказу.Зарезервируйте место и ознакомьтесь с правилами посещения!
Дайте Открыть / закрыть меню
перейти к содержанию- ВИЗИТ
- Часы, прием, бронирование
- Спланируйте посещение
- Проезд
- Что в цвету
- Садовый магазин + колода для растений
- Календарь
- О КОМПАНИИ
- Сад
- Сохранение
- Исследования
- Фауна
- Блог о саду
- Персонал и Правление
- Занятость
- Архив информационных бюллетеней
- КОЛЛЕКЦИИ
- Коллекции растений
- Тур виртуальных коллекций
- Тропический дом Проект
- ОБРАЗОВАНИЕ
- Виртуальное сообщество
- Экскурсии
- Внутришкольные программы
- Летний лагерь 2021
- Колледж и университет
- Учебная программа
- Педагоги
- ПРОГРАММЫ
- Программы и семинары
- Международные туры
- Выставок
- Семейные программы
- Летние концерты
- События
- Иллюстрированные растения 2021
- ОПОРА
- Пожертвовать
- Членство
- Волонтер
- АРЕНДА
Дайте 404
Ой!
Страница не найдена
← Перейдите на домашнюю страницу или просто выполните поиск… Искать:
- ДОМ
- ВИЗИТ | Часы работы и входной билет
- Спланируйте посещение
- Проезд
- Что в цвету
- Садовый магазин + колода для растений
- О НАС | Сад
- Сохранение
- Исследования
- Фауна
- Персонал и Правление
- Блог о саду
- Занятость
- КОЛЛЕКЦИИ
- Обзор коллекций
- ОБРАЗОВАНИЕ | Учебная программа
- Педагоги
- Внутришкольные программы
- Экскурсии
- Типы туров
- Расписание экскурсии
- Летний лагерь
- Колледж и университет
- ПРОГРАММЫ
- Программы и семинары
- События
- Выставок
- Семейные программы
- Экскурсии по саду
- Летние концерты
- ПОДДЕРЖКА | Пожертвовать
- Кирпичи дани
- Скамейки Tribute
- Усыновления деревьев
- ЧЛЕНСТВО
- Присоединяйтесь к
- Преимущества
- Питомники
- Взаимный
- FAQ
- Подарочные сертификаты и ваучеры
- Волонтер
- АРЕНДА
- Заведения
- Информация о пакете
- Тарифы и мощности
- Поставщики общественного питания
- FAQ
- Как оформить заказ
© 2020 Ботанический сад Калифорнийского университета в Беркли
Наверх ↑
. Наверх
Мы благодарим Фреда Сислинга (Ботанический сад Утрехта) за его помощь во время эксперимента и Пола Вестерса (Университетский медицинский центр Утрехта) за статистические советы. Работа поддержана Фондом Микеля (Утрехтский университет).
6″> ПРИЛОЖЕНИЕ Смертность гаметофитов в эксперименте с течением времени, усредненная по уровню инбридинга. I — интрагаметофитное самоопыление; II, внутриплощадочные кроссы; III, межузельные кресты.
© Автор, 2010. Опубликовано издательством Oxford University Press от имени компании Annals of Botany. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
. Жизненный цикл папоротника: размножение растений без цветов и семян — класс CLEP [видео 2021]
Обзор: чередование поколений
Это чередование поколений представляет собой жизненный цикл, который включает как диплоидные, так и гаплоидные многоклеточные стадии.Помните, что диплоид означает два набора хромосом и обычно обозначается сокращенно как «2N», потому что N обозначает хромосомы. В диплоидных клетках по одной копии хромосом от каждого родителя. Например, у людей вы получаете одну копию хромосом от отца и одну копию хромосом от мамы.
Та же идея встречается и в растениях. Каждая диплоидная клетка содержит одну копию хромосом от мужского родителя и одну копию хромосом от женского родителя. Гаплоид означает один набор хромосом и обычно обозначается аббревиатурой «N», потому что существует только одна копия хромосом.
Диаграмма смены поколений
Рассмотрим схему основной идеи смены поколений. На этой диаграмме видно, что жизненный цикл разбит на N вверху и 2N внизу. Помните, что гаметофит содержит гаплоидные клетки, а спорофит — диплоидные клетки. Ранее мы использовали garblinx, чтобы проиллюстрировать странность этого. Помните, что диплоидный, или 2N, организм выглядит так.
Диаграмма, показывающая диплоидный организм
Однако, когда он переходит в гаплоидную стадию, гарблинкс выглядит совершенно иначе! Два из этих гаплоидных организмов собираются вместе и спариваются, чтобы произвести новый организм 2N, похожий на первый гарблинкс. Затем этот гарблинкс в конечном итоге произведет новый гаплоидный организм и так далее.
Теперь, когда мы рассмотрели основные аспекты этого жизненного цикла, давайте посмотрим, как папоротники проходят этот цикл.Мы начнем с гаплоидной стадии, как делали раньше, а затем перейдем к стадии доминантного диплоида.
Гаплоидная стадия
Мы будем использовать эту диаграмму, чтобы проиллюстрировать жизненный цикл папоротников.
Схема жизненного цикла папоротника
Папоротник начинается с гаплоидной стадии в виде спор. Спора подвергнется митозу, чтобы создать множество идентичных гаплоидных клеток. Эта группа клеток называется гаметофитом .На нашей диаграмме видно, что спора перерастает в гаметофит. Большинство гаметофитов папоротника выглядят как листья в форме сердца и меньше вашего мизинца.
Интересно, что этот гаметофит, несмотря на невысокие размеры и структуру, способен к фотосинтезу. Это означает, что гаметофит может вырабатывать собственную энергию и расти независимо.
После развития и роста в качестве гаметофита спора готова производить гамет . Гаметы образуются в определенной области гаметофита. Gametangia — это органы гаметофитов, производящие гаметы.
Есть два типа гаметангий: один производит яйцеклетки, а другой — сперматозоиды. архегонии — это женские гаметангии, которые производят одно яйцо за раз. Антеридии — мужские гаметангии, которые производят много сперматозоидов. Вы можете вспомнить, какие гаметангии мужские, а какие женские, потому что пыльники — это мужские структуры в цветках, производящие сперму. Если вы знаете, что пыльники производят сперму, вы можете помнить, что антеридии также производят сперму и, следовательно, являются мужскими.
На нашей диаграмме показаны мужские и женские структуры одного и того же гаметофита. Это означает, что гаметофит папоротника бисексуален, потому что он содержит как мужские, так и женские структуры. Антеридии расположены ближе к верхушке сердцевидного листа. Архегонии расположены в верхней части гаметофита. С генетической точки зрения, для гамет лучше всего смешиваться между разными гаметофитами, чтобы начать диплоидную стадию. Если гаметы происходят от одного и того же растения, генетического разнообразия не будет.Однако, если яйцеклетка и сперма происходят от разных растений, это увеличит генетическую изменчивость, что, скорее всего, повысит шансы растения на выживание.
Диплоидная стадия
Диплоидная — и доминирующая — стадия жизни начнется после того, как произойдет оплодотворение. Помните, что оплодотворение — это слияние яйцеклетки и спермы. И яйцеклетка, и сперматозоид гаплоидны, поэтому, когда они сливаются вместе, мы получаем диплоидную клетку. Эта новая клетка будет продолжать митоз и расти. В конце концов, он станет зрелым спорофитом, похожим на известные вам папоротники.У зрелого спорофита много разных структур, но мы остановимся на тех, которые участвуют в размножении.
На нижней стороне листьев папоротника вы можете найти красные или коричневые точки. Эти точки называются сори и представляют собой группы спорангиев.
Что такое спорангии? Подобно тому, как у гаметофитов есть гаметангии, производящие гаметы, у спорофитов есть спорангии, производящие споры. Спорангии — это органы, производящие споры в процессе мейоза.
Фото сори на нижней стороне листьев папоротника
Внутри каждого спорангия есть клетки, которые фактически делятся и становятся спорами. Эти спороцитов представляют собой диплоидные клетки, которые подвергаются мейозу с образованием гаплоидных спор. Эти споры микроскопические, то есть очень маленькие. Хотя вы можете увидеть группы спорангиев на нижней стороне листьев папоротника, вы не можете увидеть споры без помощи микроскопа.После высвобождения эти споры начнут гаплоидную стадию жизненного цикла.
Резюме урока
Папоротники — уникальные растения, потому что они не дают семян. Эти сосудистые растения адаптировались к жизненному циклу, в котором используются споры, а не семена. Это чередование поколений у папоротников означает, что есть многоклеточные стадии, которые бывают гаплоидными и диплоидными.
Сначала мы рассмотрели гаплоидную стадию, которая начинается как спора и развивается в гаметофит .Гаметофит папоротника выглядит как небольшой лист в форме сердца и является обоеполым, он содержит как мужские, так и женские органы, производящие гамет. гаметангии , производящие сперму, называются антеридиями , а те, которые производят яйцеклетки, называются архегониями .
Как только яйцеклетка оплодотворяется спермой, папоротник переходит в диплоидную стадию жизненного цикла. Эта новая диплоидная клетка развивается в зрелый спорофит, который представляет собой структуру, о которой вы думаете, когда думаете о папоротниках.У спорофита есть уникальные структуры, называемые спорангии , расположенные на нижней стороне некоторых листьев. Эти структуры содержат спороцита , которые продуцируют споры . Эти споры снова начинают гаплоидную стадию.
Результат обучения
В конце этого урока вы сможете описать этапы уникального жизненного цикла папоротников.
Яванский папоротник: полное руководство по уходу (виды, посадка и размножение)
Яванский папоротник, с научной точки зрения известный как Microsorum pteropus , является классическим и очень популярным аквариумным растением.
Этот нежный на вид папоротник придает аквариуму красивый вид. Он широко используется из-за медленного роста, уникальной формы и простоты воспроизводства и ухода.
Может содержаться с самыми разными рыбами и не требует строгих параметров воды, поэтому так нравится многим людям. Это одно из самых простых растений для начинающих.
В этой статье мы расскажем о происхождении и внешнем виде растения, о том, как его сажать и поддерживать, о методах размножения и подходящих помощниках по аквариуму.Прежде чем продолжить чтение, вы можете взглянуть на это краткое резюме.
Категория Рейтинг Семья: Polypodiaceae Уровень ухода: Easy Скорость роста до 13,5 дюймов Минимальный размер бака: 10 галлонов Условия воды: 68-82 o F, pH 6.0-7,5, KH 3-8 Освещение: От слабого до умеренного Размножение: Придаточное растение / корневище Размещение: Середина к фону Яванский папоротник — член семейства полиподиевых, насчитывающего более 60 различных родов. Microsorum — это род этого семейства, которое включает более 50 видов тропических папоротников.
Пожалуй, одним из самых популярных видов рода Microsorum является Microsorum pteropus, также известный как Яванский папоротник.
Произошло из Юго-Восточной Азии и представляет собой растение в джунглях, которое обычно растет на камнях, земле и вокруг грузовиков с деревьями вдоль ватерлинии паров и водопадов. Он также растет в тропических дождевых лесах, как трава. Он может расти как при полностью погруженном, так и частично погруженном в воду.
Ява Папоротник давно является хобби, и с годами были выведены новые разновидности.Самые популярные из них, которые вы найдете в рыбных магазинах, — это узколистный, игольчатый, трезубец и винделов.
Это относительно дешево; Вы можете купить небольшое растение примерно за 4-5 долларов. Иногда они уже прикреплены к корягу, что позволяет легко попасть прямо в аквариум. Растения, которые уже растут на древесине, как правило, стоят дороже, в зависимости от размера, вы можете рассчитывать заплатить более 20 долларов.
Когда вы покупаете их, они обычно будут около 3-5 дюймов в длину. Убедитесь, что вы выбрали здоровое растение без коричневой окантовки и зеленых, здоровых на вид листьев.
Не покупайте пачку, иногда магазины просто собирают вместе партию черенков, и у них отсутствуют корневища.
Это отличное растение, если у вас ограниченный бюджет. Практически невозможно убить это растение, если вы не предпринимаете активных действий!
Внешний вид папоротника Явы
Папоротник Явы — традиционное зеленое аквариумное растение, состоящее из двух основных компонентов; корневище и листья.
Корневища действуют как якорь и представляют собой темно-коричневые, похожие на волоски струны, которые прикрепляются ко многим различным поверхностям.
Листья очень выносливые и имеют кожистую текстуру различных уникальных форм, от кустистых до колючих. Он бывает зеленого цвета от среднего до темно-зеленого. Обычно чем выше освещение, тем темнее зеленый цвет. На некоторых зрелых листьях появляется несколько черных / коричневых крошечных круглых бугорков (один из способов размножения, который мы вскоре обсудим), а иногда и несколько черных прожилок, проходящих через листья.
Растение может достигать высоты около 13,5 дюймов и может достигать 6-8 дюймов в ширину.Это делает его идеальным для самых разных аквариумов, от больших общественных аквариумов до густо засаженных или даже демонстрационных.
У оригинального растения есть варианты, которые влияют на размер и форму листьев. Мы рассмотрим некоторые из самых популярных разновидностей аквариумов; однако есть и другие, менее известные разновидности.
- Узколистный Яванский папоротник: У этого растения узкие листья, которые растут под более крутым углом, чем у обычного Microsorum pteropus .Листья вырастают примерно до 4-8 дюймов, а растение может вырасти до 12 дюймов.
- Игольчатый папоротник Ява: Это растение имеет еще более тонкие листья и меньше по размеру, чем узколистное растение. Он может вырасти до 6 дюймов в высоту. Настоящий игольчатый лист Ява довольно редко встречается в торговле.
- Трезубец Ява Папоротник: Это один из наиболее необычных вариантов, у него перистые лопастные листья с 2-5 долями с каждой стороны. Он короче узкого листа, но растет быстрее и имеет больше вилок на каждом листе.
- Windelov Ява Папоротник: Уникальный вариант с мелко разветвленными кончиками листьев. Обычно он вырастает примерно до 8 дюймов в высоту.
Java Fern Care
Microsorum pteropus Требования к аквариуму
Как и рыбы, водные растения процветают, когда условия аквариума максимально соответствуют их естественным условиям.
В своем естественном климате папоротник Явы растет вдоль ручьев и часто встречается у движущихся водоемов. Фильтры и насадки обеспечивают достаточное количество кислорода для этих растений в аквариуме — им не нужно много дополнительного CO2.
Их также можно найти на дне джунглей, где мягкая кислая вода брызгает на листья из ручьев и водопадов. Для воспроизведения этих условий рекомендуется значение pH от 6,0 до 7,0 с жесткостью 3-8dGH.
Это чрезвычайно выносливое растение, которому не нужны особые условия для роста. Ему не требуется яркое освещение, хороший субстрат или большое количество питательных веществ, поэтому это растение — мечта.
Чтобы он рос, вам не нужен красивый аквариум; он будет расти при самых простых настройках.По размеру его можно выращивать в резервуарах размером до 10 галлонов. Мы не рекомендуем помещать его в емкости меньшего размера. Он будет расти в большинстве условий освещения; наиболее предпочтительны приглушенные люминесцентные лампы или лампы накаливания. Не делайте освещение слишком сильным — они станут коричневыми и прозрачными, если освещение будет слишком сильным.
В этом случае уменьшите освещение или выключите его на несколько дней, пока растение не восстановится. В дикой природе эти растения фактически растут в тени и не подвергаются воздействию яркого света.
В идеале, при использовании ламп 5000-7000K на каждый галлон в вашем резервуаре требуется 1,5 / 2 Вт света.
С точки зрения температуры, растение будет расти в холодной воде и в тропических резервуарах, его предпочтительная температура составляет 68-82 o F.
Он также идеально подходит для резервуаров с голым дном, поскольку не требует никакого субстрата. Если вы все же хотите добавить субстрат в аквариум с растением, выберите все, что вам нравится; это растение не извлекает питательные вещества из субстрата.
Интересно, что некоторые люди настаивают на том, что это растение может выжить в солоноватой воде, и пока оно будет жить какое-то время, оно, скорее всего, не будет расти и в конечном итоге умрет.
Как сажать Явский папоротник
Явский папоротник, привязанный к дереву Явский папоротник имеет корневища, которые не любят закапывать. Растение будет расти очень медленно или может вообще не расти, если вы его закопаете.
Вместо того, чтобы закапывать его, вам нужно прикрепить его к корягу, камням или чему-то подобному.
Шероховатые поверхности лучше гладкие. Все, что угодно, например, лавовый камень или коряги, подойдет идеально. Если поверхность объекта слишком гладкая (например, галька или стекло), потребуется больше времени для прикрепления, но обычно он все равно в конечном итоге прикрепляется.Желательно использовать большой кусок коряги, чтобы у него было достаточно места для разложения, но можно использовать и маленькие кусочки.
Чтобы обезопасить растение, привяжите корни с помощью рыболовной проволоки к выбранному вами объекту. Черный цвет предпочтительнее использовать, так как он соответствует цвету корней. Некоторые люди предпочитают использовать резинки или стяжки, однако они обычно видны снаружи вашего бака.
Вы обнаружите, что через несколько недель корни прикрепятся к поверхности того, к чему вы их прикрепили.
Как только корни закреплены, вы можете удалить их стяжки; вы можете сделать это, если выбрали хорошо заметный галстук. Вам не нужно снимать леску и т. Д., Если вы не хотите, так как растение будет прикрывать ее по мере роста.
Яванский папоротник может вырасти довольно большими и с особенно широкими листьями; Поэтому рекомендуется сажать его посередине или ближе к задней части резервуара, если вы не пытаетесь добиться эффекта джунглей.
Если вы поместите его в передней части резервуара, он скроет все остальное, что у вас там есть.
Вы можете сажать его отдельно, группами или другими растениями.
Вместо этого вы можете захотеть поплавать это растение, однако его корневища будут расти все дольше и дольше, пока оно не найдет, к чему прикрепиться.
Уход и уход
Папоротник Ява — одно из самых простых в уходе растений; Если у вас когда-либо возникали проблемы с выращиванием аквариумных растений, попробуйте это.
Вы можете обнаружить, что вашему растению нужно время, чтобы начать расти, возможно, оно просто приспосабливается к условиям вашего аквариума и закладывает корневища.Как только это произойдет, он начнет расти.
То, как вы ухаживаете за своим, зависит от того, какой внешний вид вы хотите достичь в своем аквариуме. Если вы хотите, чтобы ваши растения были маленькими и отдельными, вам нужно будет удалить маленькие новые ростки, которые развиваются на листьях по мере их появления (мы расскажем, как это сделать в разделе о размножении).
Если вы хотите создать действительно густой рост, просто оставьте растение, чтобы оно продолжало расти. По мере развития новых проростков вместо того, чтобы удалять их для создания новых растений, оставьте их расти, чтобы добиться эффекта большой густоты.
Эти растения получают питательные вещества из воды через листья, поэтому удобрения не требуются. Однако, если вы хотите использовать удобрения для ускорения роста, вы можете это сделать. Не имея настоящих корней, они получают большую часть питательных веществ через листья из водной толщи, поэтому удобрения для субстрата будут бесполезны. Вместо этого добавляйте жидкое удобрение каждый раз при подмене воды.
Если на вашем растении появились ожоги, вы можете обрезать растение, чтобы удалить их.
Из-за медленного роста вы можете обнаружить, что вам нужно делать только одну большую обрезку в год.Обрежьте листья как можно ближе к корневищу.
Прелесть Java Fern, прикрепляющегося к камням и корягам, заключается в том, что когда дело доходит до очистки аквариума, вы можете перемещать его во время очистки. Это особенно привлекательно для аквариумов, содержащих грязную рыбу.
N.B. Когда вы чистите свой аквариум или проводите какое-либо другое обслуживание, это растение необходимо постоянно поддерживать во влажном состоянии, поэтому либо держите его под водой, либо используйте распылитель, чтобы листья оставались влажными.
Общие проблемы
Многие новички думают, что все растения нужно сажать в субстрат.Это распространенная ошибка, которая приведет к смерти Java Fern. Вы обнаружите, что он выживет в течение нескольких недель в субстрате, но затем он начнет становиться коричневым, что является признаком того, что он умирает.
Вторая проблема, с которой сталкиваются люди, заключается в том, что они обнаруживают, что растение не растет. Отсутствие роста в первые несколько недель — это нормально. Если он все еще не растет, вы можете рассмотреть возможность использования удобрений, как указано выше.
Другая распространенная ошибка, которую люди совершают с этим растением, — это принимать черные / коричневые пятна на листьях за нездоровое растение.Эти черные круги — то место, откуда будут расти новые ростки, так что не волнуйтесь! Черные пятна, которые не развиваются в новые растения, могут быть признаком дефицита азота; другие пятна могут указывать на ожоги, поэтому выключите свет.
Последняя проблема, которую мы рассмотрим, — это плавление Java Fern. Для этого характерны большие коричневые пятна, которые, в свою очередь, вызывают гниение и размягчение растения.
Обычно это происходит из-за того, что растениям не хватает питательных веществ, слишком много света или слишком много сине-зеленых водорослей в вашем аквариуме.
В качестве примечания: если у вас есть немного водорослей, вы можете подумать о том, чтобы нанять бригаду по уборке, которая поможет держать их в курсе.
Ничто из этого не должно быть проблемой, если вы правильно настроили резервуар, правильно следуете приведенному выше разделу по посадке и уходу и регулярно меняете воду.
Размножение папоротника на Яве
Фото ictheostega. Размножение Java Fern очень просто и не требует каких-либо особых условий или работы с вашей стороны.
Есть несколько способов размножения:
- Во-первых, вы можете разрезать корневище пополам и заново засеять срезанные секции.Затем отдельные растения продолжат расти.
- В качестве альтернативы можно подождать, пока на листьях растения разовьются крошечные папоротники Явы. Иногда они начинаются с черных пятен на нижней стороне листа.
Примерно через 2-3 недели из черных шишек начинают прорастать крошечные листья.
Их можно легко срезать острыми стерилизованными ножницами и переместить в другое место на растении, чтобы дать начало новому росту. По мере развития и созревания ростки снова начинают процесс роста.
Помните, что новый завод все равно нужно будет привязать так же, как мы обсуждали ранее.
Соседи и совместимость с папоротником Java в аквариуме
Рыба-ангел в аквариуме с растениями Это растение очень совместимо с огромным разнообразием рыб.
Даже травоядные рыбы, обычно питающиеся аквариумными растениями, стараются держаться от них подальше. Он также имеет очень жесткую структуру листьев, что делает его непривлекательным для рыбы.
Он совместим как с игривыми мелкими рыбками, так и с крупными, такими как цихлиды, тигровые оскары, сомы, арованы и другие крупные рыбы, которые обычно повреждают более чувствительные растения.
Однако делайте это с осторожностью в начале жизненного цикла растений. Его корни еще не так прочны; следовательно, он будет подвержен повреждениям и опрокидыванию, если вы решите разместить в нем крупную и более агрессивную рыбу.
Если вы все-таки живете с более крупной рыбой, прикрепите растение к корягу или камням, чтобы не повредить корни.
Яванский папоротник — очень выносливое, сильное и крепкое растение, которое трудно разорвать на части. Если вы хотите, чтобы растение выглядело гладким и красивым, вам не следует размещать его вместе с игривыми или мелкими рыбками.
Подходит ли Яванский папоротник для вашего аквариума (резюме)?
Если у вас пресноводный аквариум, то почти 100% вероятность, что это растение подойдет ему.
Если вы новичок в этом хобби или опытный ветеран, это растение обладает множеством замечательных качеств, которые делают его желанным аквариумным растением.
За ним очень легко ухаживать и поддерживать. Его легко размножать, и с ним можно содержать широкий спектр рыб.
Доступно несколько различных вариантов; тип, который вы выберете, будет зависеть от наличия и личных предпочтений.
Есть ли у вас опыт работы с Java Fern? Мы хотели бы услышать — напишите нам сообщение в разделе комментариев ниже…
Уникальная система соматического эмбриогенеза в древовидном папоротнике Cyathea delgadii Sternb: важность типа эксплантата и физических и химических факторов
Akula A, Becker D, Bateson M (2000) Высокоурожайный повторяющийся соматический эмбриогенез и восстановление растений в выбранном чайном клоне TRI-2025 путем временного погружения. Plant Cell Rep 19: 1140–1145
CAS
Статья
Google Scholar
Ambrósio ST, de Melo NF (2004) Взаимодействие между сахарозой и pH во время культивирования in vitro Nephrolepis biserrata (Sw.) Шотт (Pteridophyta). Acta Bot Bras 18: 809–813
Артикул
Google Scholar
Ambrožič-Dolinšek J, Camloh M (1997) Гаметофитная и спорофитная регенерация из почковой чешуи папоротника Platycerium bifurcatum (Cav.) C. Chr. in vitro. Ann Bot 80: 23–28
Статья
Google Scholar
Bharati SK, Manabendra DC, Behari MP (2013) Размножение in vitro у Pteridophytes: обзор.Int J Res Ayurveda Pharm 4: 297–303
Статья
Google Scholar
Bui LT, Hurst A, Irish EE, Cheng CL (2012) Влияние сахаров и этилена на апоспорию и регенерацию у Ceratopteris richardii . Am J Plant Sci 3: 953–961
CAS
Статья
Google Scholar
Chen J-T, Chang W-C (2002) Влияние условий культивирования тканей и характеристик эксплантов на прямой соматический эмбриогенез у Oncidium «Gower Ramsey».Культ растительных клеток, тканей и органов 69: 41–44
CAS
Статья
Google Scholar
Cordle AR, Irish EE, Cheng C-L (2007) Индукция апогамии у Ceratopteris richardii . Int J Plant Sci 168: 361–369
Статья
Google Scholar
Cordle AR, Bui LT, Irish EE, Cheng CL (2011) Апогамия и апоспория, вызванные лабораторными методами, у Ceratopteris richardii .В: Фернандес Х., Кумар А., Ревилла М.А. (ред.) Работа с папоротниками: проблемы и приложения. Springer Science + Business Media, LLC, Берлин, стр. 25–36
Google Scholar
Cunha A, Fernandes-Ferreira M (1999) Влияние параметров среды на соматический эмбриогенез из эксплантатов гипокотилей льна ( Linum usitatissimum L.). J Plant Physiol 155: 591–597
CAS
Статья
Google Scholar
Dykeman BW, Cumming BG (1985) Размножение страусиного папоротника in vitro ( Matteuccia struthiopteris ).Can J Plant Sci 65: 1025–1032
CAS
Статья
Google Scholar
Fernández H, Revilla MA (2003) In vitro Культивирование декоративных папоротников. Культ растительных клеток, тканей и органов 73: 1–13
Статья
Google Scholar
Фернандес Х., Бертран А.М., Санчес-Тамес Р. (1996) Влияние условий культивирования тканей на апогамию у Driopteris affinis sp. affinis . Культ растительных клеток, тканей и органов 45: 93–97
Статья
Google Scholar
Фернандес Х., Бертран А.М., Санчес-Тамес Р. (1999) Биологические и пищевые аспекты, связанные с размножением папоротников. Культ растительных клеток, тканей и органов 56: 211–214
Статья
Google Scholar
Ferreira DAT, Sattler MC, Carvalho CR, Clarindo WR (2015) Эмбриогенный потенциал незрелых зиготических зародышей Passiflora : новое достижение для размножения in vitro без регуляторов роста растений.Культ органа растительной клетки, ткани. DOI: 10.1007 / s11240-015-0796-1
Google Scholar
Филонова Л.Х., Божков П.В., фон Арнольд С. (2000) Путь развития соматического эмбриогенеза у Picea abies по данным покадровой съемки. J Exp Bot 51: 249–264
CAS
Статья
PubMed
Google Scholar
Gaj M (2004) Факторы, влияющие на индукцию соматического эмбриогенеза и регенерацию растений с особым упором на Arabidopsis thaliana (L.) Heynh. Регламент роста растений 43: 27–47
CAS
Статья
Google Scholar
Gow W-P, Chen J-T, Chang W-C (2010) Усиление прямого соматического эмбриогенеза и роста ростков из листовых эксплантатов Phalaenopsis путем регулирования периода культивирования и длины эксплантата. Acta Physiol Plant 32: 621–627
Статья
Google Scholar
Gray DJ, McColley DW, Compton ME (1993) Высокочастотный соматический эмбриогенез из покоящихся семядолей семян сортов Cucumis melo .J Am Soc Hort Sci 118: 425–432
Google Scholar
Hirsch AM (1975) Влияние сахарозы на дифференциацию вырезанной ткани листа папоротника на гаметофиты или спорофиты. Физиология растений 56: 390–393
PubMed Central
CAS
Статья
PubMed
Google Scholar
Hirsch AM (1976) Развитие апоспоровых гаметофитов и регенерированных спорофитов из эпидермальных клеток иссеченных листьев папоротника: анатомическое исследование.Am J Bot 63: 263–271
Статья
Google Scholar
Хименес В.М. (2005) Участие растительных гормонов и регуляторов роста растений в соматическом эмбриогенезе in vitro. Регламент роста растений 47: 91–110
Статья
Google Scholar
Камада Х., Кобаяши К., Кийосуэ Т., Харада Х. (1989) Стресс-индуцированный соматический эмбриогенез в моркови и его применение для производства синтетических семян.In vitro Cell Dev Biol 25: 1163–1166
Статья
Google Scholar
Камада Х, Исикава К., Сага Х, Харада Х (1993) Индукция соматического эмбриогенеза в моркови под действием осмотического стресса. Plant Tissue Cult Lett 10: 38–44
CAS
Статья
Google Scholar
Karami O, Deljou A, Esna-Ashari M, Ostad-Ahmadi P (2006) Влияние концентраций сахарозы на соматический эмбриогенез у гвоздики ( Dianthus caryophyllus L.). Sci Hortic 110: 340–344
CAS
Статья
Google Scholar
Курияма А., Ходжох Т., Сугавара Ю., Мацусима Х., Такеучи М. (1989) Метод быстрого роста в культуре гаметофитов Equisetum arvense с антеридиями. Физиология растительных клеток 30: 1189–1192
Google Scholar
Курияма А., Кобаяси Т., Маеда М. (2004) Получение спорофитных растений Cyathea lepifera , древовидного папоротника, из гаметофита, культивируемого in vitro.J Jpn Soc Hortic Sci 73: 140–142
Статья
Google Scholar
Liao YK, Wu YH (2011) In vitro размножение Platycerium bifurcatum (Cav.) C. Chr. через инициирование зеленого шаровидного тела. Bot Stud 52: 455–463
CAS
Google Scholar
Лу Х., Како С. (1995) Роль высоких концентраций сахара в индукции соматического эмбриогенеза семядолей огурца.Sci Hortic 64: 11–20
CAS
Статья
Google Scholar
Mahdavi-Darvari F, Mohd Noor N, Ismanizan I (2015) Эпигенетическая регуляция и генные маркеры как сигналы раннего соматического эмбриогенеза. Культ растительных клеток, тканей и органов 120: 407–422
CAS
Статья
Google Scholar
Martin KP, Sini S, Zhang C-L, Slater A, Madhusoodanan PV (2006) Эффективная индукция апоспории и апогамии in vitro в серебряном папоротнике ( Pityrogramma calomelanos L.). Plant Cell Rep 25: 1300–1307
CAS
Статья
PubMed
Google Scholar
Materi DM, Cumming BG (1991) Влияние углеводной депривации на омоложение, апоспорию и регенерацию у спорофитов страусиного папоротника ( Matteuccia struthiopteris ). Can J Bot 69: 1241–1245
Статья
Google Scholar
Menéndez V, Villacorta NF, Revilla MA, Gotor V, Bernard P, Fernández H (2006) Экзогенные и эндогенные регуляторы роста при апогамии у Dryopteris affinis (Lowe) Fraser-Jenkins sp. affinis . Plant Cell Rep 25: 85–91
Статья
PubMed
Google Scholar
Mikuła A, Pożoga M, Tomiczak K, Rybczyński J (2015) Соматический эмбриогенез в папоротниках: новая экспериментальная система. Plant Cell Rep 34: 783–794
PubMed Central
Статья
PubMed
Google Scholar
Moura IR, Simões-Costa MC, Garcia J, Silva MJ, Duarte MC (2012) Культивирование in vitro спор древесных папоротников из семейств Cyatheaceae и Dicksoniaceae .Acta Hortic 937: 455–461
Статья
Google Scholar
Murashige T, Skoog F (1962) Пересмотренная среда для быстрого роста и биоанализов с культурами тканей табака. Physiol Plant 15: 473–497
CAS
Статья
Google Scholar
Nakagawa H, Saijyo T, Yamauchi N, Shigyo M, Kako S, Ito A (2001) Влияние сахаров и абсцизовой кислоты на соматический эмбриогенез из мело ( Cucumis melo L.) расширенная семядоля. Sci Hortic 90: 85–92
CAS
Статья
Google Scholar
Nhut DT, Le BV, Fukai S, Tanaka M, Van KTT (2001) Влияние активированного угля, размера эксплантата, положения экспланта и концентрации сахарозы на регенерацию растений и побегов Lilium longiflorum через культивирование молодых стеблей. Регламент роста растений 33: 59–65
Статья
Google Scholar
Parajuli J, Joshi SD (2014) In vitro исследование эффектов гормонов роста на развитие спорофитов Cyathea spinulosa .Int J Biodivers Conserv 6: 247–255
Статья
Google Scholar
Pinto G, Park Y-S, Silva S, Neves L, Araújo C, Santos C (2008) Факторы, влияющие на поддержание, пролиферацию и прорастание вторичных соматических зародышей Eucalyptus globulus Labill. Культ организации тканей клеток растений 95: 69–78
CAS
Статья
Google Scholar
Raemakers CJJM, Jacobsen E, Visser RGF (1995) Вторичный соматический эмбриогенез и применение в селекции растений.Euphytica 81: 93–107
Статья
Google Scholar
Рагхаван В. (1989) Биология развития гаметофитов папоротников. Cambridge University Press, Cambridge, pp. 259–295
Книга
Google Scholar
Рашид А. (2009) Введение в мохообразные: разнообразие, развитие и дифференциация. В: Глава 11: смена поколений. VIKAS Publishing House PVT LTD, Нью-Дели, стр. 237–249
Рыбчинский Ю., Микула А. (2011) Биотехнология древовидных папоротников: от спор до спорофитов.В: Фернандес Х., Кумар А., Ревилла М.А. (ред.) Работа с папоротниками: проблемы и приложения. Springer Science + Business Media, Нью-Йорк, Дордрехт, Гейдельберг, Лондон, стр 135–147
Google Scholar
Смит Д.Л., Крикориан А.Д. (1990) Производство соматических проэмбрионов из иссеченных, раненых зародышей зиготической моркови на среде без гормонов: оценка влияния pH, этилена и активированного угля. Plant Cell Rep 9: 34–37
CAS
Статья
Google Scholar
Somer A, Arbesú R, Menéndez V, Revilla MA, Fernández H (2010) Исследования индукции спорофитов в папоротниках in vitro.Euphytica 171: 203–210
Статья
Google Scholar
Такамура Т., Танака М. (1996) Соматический эмбриогенез из этиолированного черешка цикламена ( Cyclamen persicum Mill.). Plant Tissue Lett 13: 43–48
CAS
Статья
Google Scholar
Tavares JE, Sussex IM (1968) Расширенный рост изолированных листовых пластинок Todea barbara .Planta 80: 113–128
CAS
Статья
Google Scholar
White RA (1971) Экспериментальные и эволюционные исследования спорофита папоротника. Bot Rev 37: 509–540
Статья
Google Scholar
Whittier DP, Pratt LH (1971) Влияние качества света на индукцию апогамии в отростках Pteridium aquilinum . Planta 99: 174–178
CAS
Статья
PubMed
Google Scholar
Whittier DP, Steeves TA (1960) Вызвание апогамии у папоротника папоротника.Can J Bot 38: 925–930
CAS
Статья
Google Scholar
Ян X, Чжан X (2010) Регулирование соматического эмбриогенеза у высших растений. Crit Rev Plant Sci 29: 36–57
CAS
Статья
Google Scholar
Ян-Лин С., Сун-Кван Х. (2012) Последние достижения в области эмбриогенеза однодольных и двудольных растений in vitro. В: Кен-Ичи Сато (ред) Эмбриогенез.ISBN: 978-953-51-0466-7, InTech. http://www.intechopen.com/books/embryogenesis/recent-advances-of-invitro-embryogenesis-of-monocotyledon-and-dicotyledon
Заваттьери М.А., Фредерико А.М., Лима М., Сабино Р., Арнхольдт-Шмитт Б. (2010) Индукция соматического эмбриогенеза как пример стресс-связанных реакций растений. Electron J Biotech 13. http://www.ejbiotechnology.cl/content/vol13/issue1/full/4/
Страница не найдена — Ботанический сад Калифорнийского университета
Сад открыт только по предварительному заказу.Зарезервируйте место и ознакомьтесь с правилами посещения!
Дайте Открыть / закрыть меню
перейти к содержанию- ВИЗИТ
- Часы, прием, бронирование
- Спланируйте посещение
- Проезд
- Что в цвету
- Садовый магазин + колода для растений
- Календарь
- О КОМПАНИИ
- Сад
- Сохранение
- Исследования
- Фауна
- Блог о саду
- Персонал и Правление
- Занятость
- Архив информационных бюллетеней
- КОЛЛЕКЦИИ
- Коллекции растений
- Тур виртуальных коллекций
- Тропический дом Проект
- ОБРАЗОВАНИЕ
- Виртуальное сообщество
- Экскурсии
- Внутришкольные программы
- Летний лагерь 2021
- Колледж и университет
- Учебная программа
- Педагоги
- ПРОГРАММЫ
- Программы и семинары
- Международные туры
- Выставок
- Семейные программы
- Летние концерты
- События
- Иллюстрированные растения 2021
- ОПОРА
- Пожертвовать
- Членство
- Волонтер
- АРЕНДА
Дайте 404
Ой!
Страница не найдена
← Перейдите на домашнюю страницу или просто выполните поиск… Искать:
- ДОМ
- ВИЗИТ | Часы работы и входной билет
- Спланируйте посещение
- Проезд
- Что в цвету
- Садовый магазин + колода для растений
- О НАС | Сад
- Сохранение
- Исследования
- Фауна
- Персонал и Правление
- Блог о саду
- Занятость
- КОЛЛЕКЦИИ
- Обзор коллекций
- ОБРАЗОВАНИЕ | Учебная программа
- Педагоги
- Внутришкольные программы
- Экскурсии
- Типы туров
- Расписание экскурсии
- Летний лагерь
- Колледж и университет
- ПРОГРАММЫ
- Программы и семинары
- События
- Выставок
- Семейные программы
- Экскурсии по саду
- Летние концерты
- ПОДДЕРЖКА | Пожертвовать
- Кирпичи дани
- Скамейки Tribute
- Усыновления деревьев
- ЧЛЕНСТВО
- Присоединяйтесь к
- Преимущества
- Питомники
- Взаимный
- FAQ
- Подарочные сертификаты и ваучеры
- Волонтер
- АРЕНДА
- Заведения
- Информация о пакете
- Тарифы и мощности
- Поставщики общественного питания
- FAQ
- Как оформить заказ
© 2020 Ботанический сад Калифорнийского университета в Беркли
Наверх ↑
. Наверх
Смертность гаметофитов в эксперименте с течением времени, усредненная по уровню инбридинга. I — интрагаметофитное самоопыление; II, внутриплощадочные кроссы; III, межузельные кресты.
© Автор, 2010. Опубликовано издательством Oxford University Press от имени компании Annals of Botany. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
.Жизненный цикл папоротника: размножение растений без цветов и семян — класс CLEP [видео 2021]
Обзор: чередование поколений
Это чередование поколений представляет собой жизненный цикл, который включает как диплоидные, так и гаплоидные многоклеточные стадии.Помните, что диплоид означает два набора хромосом и обычно обозначается сокращенно как «2N», потому что N обозначает хромосомы. В диплоидных клетках по одной копии хромосом от каждого родителя. Например, у людей вы получаете одну копию хромосом от отца и одну копию хромосом от мамы.
Та же идея встречается и в растениях. Каждая диплоидная клетка содержит одну копию хромосом от мужского родителя и одну копию хромосом от женского родителя. Гаплоид означает один набор хромосом и обычно обозначается аббревиатурой «N», потому что существует только одна копия хромосом.
Рассмотрим схему основной идеи смены поколений. На этой диаграмме видно, что жизненный цикл разбит на N вверху и 2N внизу. Помните, что гаметофит содержит гаплоидные клетки, а спорофит — диплоидные клетки. Ранее мы использовали garblinx, чтобы проиллюстрировать странность этого. Помните, что диплоидный, или 2N, организм выглядит так.
Однако, когда он переходит в гаплоидную стадию, гарблинкс выглядит совершенно иначе! Два из этих гаплоидных организмов собираются вместе и спариваются, чтобы произвести новый организм 2N, похожий на первый гарблинкс. Затем этот гарблинкс в конечном итоге произведет новый гаплоидный организм и так далее.
Теперь, когда мы рассмотрели основные аспекты этого жизненного цикла, давайте посмотрим, как папоротники проходят этот цикл.Мы начнем с гаплоидной стадии, как делали раньше, а затем перейдем к стадии доминантного диплоида.
Гаплоидная стадия
Мы будем использовать эту диаграмму, чтобы проиллюстрировать жизненный цикл папоротников.
Папоротник начинается с гаплоидной стадии в виде спор. Спора подвергнется митозу, чтобы создать множество идентичных гаплоидных клеток. Эта группа клеток называется гаметофитом .На нашей диаграмме видно, что спора перерастает в гаметофит. Большинство гаметофитов папоротника выглядят как листья в форме сердца и меньше вашего мизинца.
Интересно, что этот гаметофит, несмотря на невысокие размеры и структуру, способен к фотосинтезу. Это означает, что гаметофит может вырабатывать собственную энергию и расти независимо.
После развития и роста в качестве гаметофита спора готова производить гамет . Гаметы образуются в определенной области гаметофита. Gametangia — это органы гаметофитов, производящие гаметы.
Есть два типа гаметангий: один производит яйцеклетки, а другой — сперматозоиды. архегонии — это женские гаметангии, которые производят одно яйцо за раз. Антеридии — мужские гаметангии, которые производят много сперматозоидов. Вы можете вспомнить, какие гаметангии мужские, а какие женские, потому что пыльники — это мужские структуры в цветках, производящие сперму. Если вы знаете, что пыльники производят сперму, вы можете помнить, что антеридии также производят сперму и, следовательно, являются мужскими.
На нашей диаграмме показаны мужские и женские структуры одного и того же гаметофита. Это означает, что гаметофит папоротника бисексуален, потому что он содержит как мужские, так и женские структуры. Антеридии расположены ближе к верхушке сердцевидного листа. Архегонии расположены в верхней части гаметофита. С генетической точки зрения, для гамет лучше всего смешиваться между разными гаметофитами, чтобы начать диплоидную стадию. Если гаметы происходят от одного и того же растения, генетического разнообразия не будет.Однако, если яйцеклетка и сперма происходят от разных растений, это увеличит генетическую изменчивость, что, скорее всего, повысит шансы растения на выживание.
Диплоидная стадия
Диплоидная — и доминирующая — стадия жизни начнется после того, как произойдет оплодотворение. Помните, что оплодотворение — это слияние яйцеклетки и спермы. И яйцеклетка, и сперматозоид гаплоидны, поэтому, когда они сливаются вместе, мы получаем диплоидную клетку. Эта новая клетка будет продолжать митоз и расти. В конце концов, он станет зрелым спорофитом, похожим на известные вам папоротники.У зрелого спорофита много разных структур, но мы остановимся на тех, которые участвуют в размножении.
На нижней стороне листьев папоротника вы можете найти красные или коричневые точки. Эти точки называются сори и представляют собой группы спорангиев.
Что такое спорангии? Подобно тому, как у гаметофитов есть гаметангии, производящие гаметы, у спорофитов есть спорангии, производящие споры. Спорангии — это органы, производящие споры в процессе мейоза.
Внутри каждого спорангия есть клетки, которые фактически делятся и становятся спорами. Эти спороцитов представляют собой диплоидные клетки, которые подвергаются мейозу с образованием гаплоидных спор. Эти споры микроскопические, то есть очень маленькие. Хотя вы можете увидеть группы спорангиев на нижней стороне листьев папоротника, вы не можете увидеть споры без помощи микроскопа.После высвобождения эти споры начнут гаплоидную стадию жизненного цикла.
Резюме урока
Папоротники — уникальные растения, потому что они не дают семян. Эти сосудистые растения адаптировались к жизненному циклу, в котором используются споры, а не семена. Это чередование поколений у папоротников означает, что есть многоклеточные стадии, которые бывают гаплоидными и диплоидными.
Сначала мы рассмотрели гаплоидную стадию, которая начинается как спора и развивается в гаметофит .Гаметофит папоротника выглядит как небольшой лист в форме сердца и является обоеполым, он содержит как мужские, так и женские органы, производящие гамет. гаметангии , производящие сперму, называются антеридиями , а те, которые производят яйцеклетки, называются архегониями .
Как только яйцеклетка оплодотворяется спермой, папоротник переходит в диплоидную стадию жизненного цикла. Эта новая диплоидная клетка развивается в зрелый спорофит, который представляет собой структуру, о которой вы думаете, когда думаете о папоротниках.У спорофита есть уникальные структуры, называемые спорангии , расположенные на нижней стороне некоторых листьев. Эти структуры содержат спороцита , которые продуцируют споры . Эти споры снова начинают гаплоидную стадию.
Результат обучения
В конце этого урока вы сможете описать этапы уникального жизненного цикла папоротников.
Яванский папоротник: полное руководство по уходу (виды, посадка и размножение)
Яванский папоротник, с научной точки зрения известный как Microsorum pteropus , является классическим и очень популярным аквариумным растением.
Этот нежный на вид папоротник придает аквариуму красивый вид. Он широко используется из-за медленного роста, уникальной формы и простоты воспроизводства и ухода.
Может содержаться с самыми разными рыбами и не требует строгих параметров воды, поэтому так нравится многим людям. Это одно из самых простых растений для начинающих.
В этой статье мы расскажем о происхождении и внешнем виде растения, о том, как его сажать и поддерживать, о методах размножения и подходящих помощниках по аквариуму.Прежде чем продолжить чтение, вы можете взглянуть на это краткое резюме.
| Категория | Рейтинг | ||
|---|---|---|---|
| Семья: | Polypodiaceae | ||
| Уровень ухода: | Easy | ||
| Скорость роста | до | 13,5 дюймов | |
| Минимальный размер бака: | 10 галлонов | ||
| Условия воды: | 68-82 o F, pH 6.0-7,5, KH 3-8 | ||
| Освещение: | От слабого до умеренного | ||
| Размножение: | Придаточное растение / корневище | ||
| Размещение: | Середина к фону |